- Как движется вода по стеблю
- Презентация «Передвижение воды и минеральных веществ по стеблю»
- Дистанционное обучение как современный формат преподавания
- Современные педтехнологии в деятельности учителя
- Биология: теория и методика преподавания в образовательной организации
- Оставьте свой комментарий
- Подарочные сертификаты
- Передвижение воды в растениях
Как движется вода по стеблю
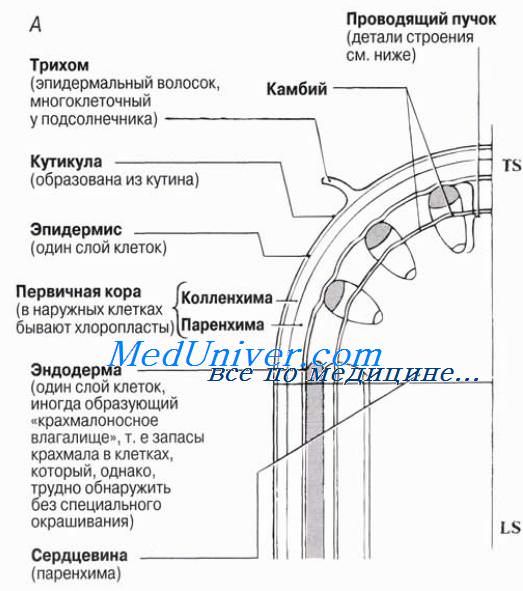
Ксилема цветковых растений содержит два типа проводящих воду структур — трахеиды и сосуды. В статье мы уже говорили о том, как выглядят эти структуры в световом микроскопе, а также на микрофотографиях, полученных с помощью сканирующего электронного микроскопа. Строение вторичной ксилемы (древесины) рассматривается в статье. Ксилема и флоэма образуют проводящую ткань высших, или сосудистых, растений. Эта ткань состоит из так называемых проводящих пучков, строение и распределение которых в стеблях двудольных растений с первичным строением показано на рисунке.
То, что вода поднимается именно по ксилеме, нетрудно продемонстрировать, погрузив побег срезанным концом в разбавленный водный раствор красителя, например эозина. Подкрашенная жидкость, распространившись вверх по стеблю, заполнит сеть пронизывающих листья жилок. Если затем сделать тонкие срезы и рассмотреть их в световом микроскопе, то окажется, что краситель находится в ксилеме.
Более эффектное доказательство подъема воды по ксилеме дают опыты с «кольцеванием». Такие опыты проводили задолго до того, как стали применяться радиоактивные изотопы, позволяющие очень легко проследить путь веществ в живом организме. В одном из вариантов опыта с одревесневшего стебля снимают узкое кольцо коры вместе с лубом, т. е. флоэмой. Довольно долго после этого находящиеся выше вырезанного кольца побеги продолжают расти нормально: следовательно, такое кольцевание не влияет на подъем воды по стеблю. Однако, если, приподняв лоскут коры, вырезать из-под него сегмент древесины, т. е. ксилемы, то растение быстро завянет. Таким образом, вода движется в побеги из почвы именно по этой проводящей ткани.
Любая теория, объясняющая транспорт воды по ксилеме, не может не учитывать следующие наблюдения.
1. Анатомические элементы ксилемы — тонкие мертвые трубки, диаметр которых варьирует от 0,01 мм в «летней» древесине до 0,2 мм в «весенней» древесине.
2. Большие количества воды движутся по ксилеме с относительно высокой скоростью: у высоких деревьев она составляет до 8 м/ч, а у других растений — около 1 м/ч.
3. Для подъема воды по таким трубкам к вершине высокого дерева необходимо давление порядка 4000 кПа. Самые высокие деревья — секвойи в Калифорнии и эвкалипты в Австралии — достигают в высоту более 100 м. Вода способна подниматься по тонким смачивающимся трубкам благодаря своему высокому поверхностному натяжению (это явление называется капиллярностью), однако только за счет этих сил даже по самым тончайшим сосудам ксилемы вода не поднимается выше 3 м.
Удовлетворительное объяснение этим фактам дает теория сцепления (когезии), или теория натяжения. Согласно этой теории, подъем воды от корней обусловлен ее испарением клетками листа. Как мы уже говорили в статье, испарение снижает водный потенциал клеток мезофилла, прилежащих к ксилеме, и вода поступает в эти клетки из ксилемного сока, водный потенциал которого выше; при этом она проходит через влажные клеточные стенки у концов жилок, как показано на рисунке.
Ксилемные сосуды заполняет сплошной столб воды; по мере того как вода выходит из сосудов, в этом столбе создается натяжение; оно передается вниз по стеблю до самого корня благодаря сцеплению (когезии) молекул воды. Эти молекулы стремятся «прилипнуть» друг к другу, потому что они полярны и притягиваются друг к другу электрическими силами, а затем удерживаются вместе водородными связями. Кроме того, они притягиваются к стенкам ксилемных сосудов, т. е. происходит их адгезия (прилипание) к ним. Сильная когезия молекул воды означает, что ее столб трудно разорвать — у него высокий предел прочности при растяжении. Растягивающее напряжение в клетках ксилемы приводит к генерированию силы, способной сдвигать весь водяной столб вверх по механизму объемного потока. Снизу вода поступает в ксилему из соседних клеток корня. При этом очень важно, что стенки ксилемных элементов жесткие и не спадаются при падении давления внутри, как это бывает, когда сосешь коктейль через мягкую соломинку. Жесткость стенок обеспечивается лигнином. Доказательством того, что жидкость внутри ксилемных сосудов сильно напряжена (растянута), служат суточные колебания диаметра древесных стволов, измеряемые инструментом под названием дендрограф.
Минимальный диаметр отмечен днем, когда интенсивность транспирации наивысшая. Натяжение столба воды в ксилемном сосуде немного втягивает внутрь его стенки (из-за адгезии), и сочетание этих микроскопических сжатий дает фиксируемую прибором общую «усадку» ствола.
Оценки прочности на разрыв столба ксилемного сока варьировали от 3000 доЗО 000 к Па, причем более низкие значения получены позднее. В листьях зарегистрирован водный потенциал порядка —4000 кПа, и прочность столба ксилемного сока, вероятно, достаточна, чтобы выдержать создающееся натяжение. Не исключено, конечно, что столб воды может иногда разрываться, особенно в сосудах большого диаметра.
Критики изложенной теории подчеркивают, что любое нарушение непрерывности столба сока должно немедленно останавливать весь поток, так как сосуд заполнится воздухом и паром (явление кавитации). Кавитацию может вызвать сильное сотрясение, изгибание ствола, а также дефицит воды. Хорошо известно, что на протяжении лета содержание воды в стволе дерева постепенно снижается, древесина заполняется воздухом. Этим пользуются лесозаготовители, потому что такие деревья легче сплавлять. Однако разрыв водного столба в части сосудов слабо влияет на общую скорость объемного потока. Возможно, дело в том, что вода перетекает в параллельно проходящие сосуды или же обходит воздушную пробку, продвигаясь по соседним паренхимным клеткам и по стенкам. Кроме того, согласно расчетам, для поддержания наблюдаемой скорости потока вполне достаточно, чтобы в каждый момент времени функционировала хотя бы небольшая доля ксилемных элементов. У некоторых деревьев и кустарников вода перемещается лишь по более молодой наружной древесине, называемой заболонью. У дуба и ясеня, например, проводящую функцию выполняют в основном сосуды текущего года, а остальная часть заболони играет роль водного резерва. Новые ксилемные сосуды образуются на протяжении всего вегетационного периода, но главным образом в его начале, когда скорость водного потока максимальна.
Вторая сила, обеспечивающая движение воды по ксилеме, — корневое давление. Его можно обнаружить и измерить в тот момент, когда срезают крону, а штамб с корнями некоторое время продолжает выделять сок из сосудов ксилемы. Этот процесс подавляется ингибиторами дыхания, например цианидом, и прекращается при недостатке кислорода и понижении температуры. Работа такого механизма, по-видимому, обусловлена активной секрецией солей и других водорастворимых веществ в ксилемный сок. В результате его водный потенциал падает, и вода поступает в ксилему из соседних клеток корня путем осмоса.
Этот механизм создает гидростатическое давление порядка 100-200 кПа (в исключительных случаях 800 кПа); одного его для подъема воды по ксилеме обычно недостаточно, однако у многих растений оно, несомненно, способствует поддержанию ксилемного тока. У медленно транспирирующих травянистых форм этого давления вполне хватает, чтобы вызвать у них мутацию. Так называется выделение воды на поверхности растения1 в виде капель жидкости, а не пара. Все условия, тормозящие транспира-цию, например слабая освещенность и высокая влажность, способствуют гуттации. Она обычна у многих видов дождевых тропических лесов и часто наблюдается на кончиках листьев у всходов трав.
Источник
Презентация «Передвижение воды и минеральных веществ по стеблю»

Описание презентации по отдельным слайдам:
Передвижение воды и минеральных веществ по стеблю Выполнил: Судак Александр 6 б класс Учитель: Полянина Ж.Л.
Стебель – осевая часть побега, он проводит питательные вещества и выносит листья к свету. В стебле могут откладываться запасные питательные вещества. На нем развиваются листья, цветки, плоды с семенами.
Для нормальной жизнедеятельности растения вода и питательные вещества должны поступать во все органы. Одна из важнейших функций стебля – транспортная. Она заключается в передаче растворов от органов почвенного питания – корней и органов воздушного питания – листьев ко всем органам растения. В этом легко убедиться, сделав продольный и поперечный срезы стебля растения.
Все растение пронизано проводящими тканями. По одним проводящим тканям движется вода с растворенными в ней минеральными веществами, по другим – раствор органических веществ. Проводящие ткани объединяются в сосудисто-волокнистые пучки, часто окруженные прочными волокнами механической ткани.
Сосудисто-волокнистые пучки проходят по всему стеблю, соединяя корневую систему с листьями. Для того, чтобы доказать это, я провел следующий опыт.
убедиться, что сосудисто-волокнистые пучки соединяют корневую систему с листьями.
1. Я сделал продольный и поперечный срезы ветки липы.
2. Я поставил веточку растения на некоторое время в подкрашенную воду. В опыте эта вода заменяет минеральные вещества. 3. Через 3 часа я сделал поперечный и продольный срезы ветки.
1.Древесина изменила свою окраску и стала синей. 2.Кора и сердцевина остались неокрашенными.
1. Растворы минеральных веществ, как и подкрашенная вода поднимаются от корня внутри стебля по сосудам древесины. 2. По этим сосудам вода с растворенными минеральными веществами и поступает в листья. Это хорошо видно на продольном и поперечном срезе стебля. 3. Большое значение для поднятия воды в стебель имеет корневое давление и испарение воды листьями. На место испарившейся воды в листья постоянно поступает новая.

Курс повышения квалификации
Дистанционное обучение как современный формат преподавания
Курс повышения квалификации
Современные педтехнологии в деятельности учителя
Курс профессиональной переподготовки
Биология: теория и методика преподавания в образовательной организации
Онлайн-конференция для учителей, репетиторов и родителей
Формирование математических способностей у детей с разными образовательными потребностями с помощью ментальной арифметики и других современных методик

Номер материала: ДВ-258855
Международная дистанционная олимпиада Осень 2021
Не нашли то что искали?
Вам будут интересны эти курсы:
Оставьте свой комментарий
Авторизуйтесь, чтобы задавать вопросы.

В России объявлены нерабочие дни с 30 октября по 7 ноября
Время чтения: 2 минуты
В московском метро появились наклейки для потерявшихся детей
Время чтения: 1 минута
В школе в Пермском крае произошла стрельба
Время чтения: 1 минута
Рособрнадзор рассчитывает, что экспресс-тесты на ковид в школах помогут избежать удаленки
Время чтения: 1 минута
Рособрнадзор оставил за регионами решение о дополнительных школьных каникулах
Время чтения: 1 минута
В России появился стандарт сбора цифрового следа в образовании
Время чтения: 2 минуты
Подарочные сертификаты
Ответственность за разрешение любых спорных моментов, касающихся самих материалов и их содержания, берут на себя пользователи, разместившие материал на сайте. Однако администрация сайта готова оказать всяческую поддержку в решении любых вопросов, связанных с работой и содержанием сайта. Если Вы заметили, что на данном сайте незаконно используются материалы, сообщите об этом администрации сайта через форму обратной связи.
Все материалы, размещенные на сайте, созданы авторами сайта либо размещены пользователями сайта и представлены на сайте исключительно для ознакомления. Авторские права на материалы принадлежат их законным авторам. Частичное или полное копирование материалов сайта без письменного разрешения администрации сайта запрещено! Мнение администрации может не совпадать с точкой зрения авторов.
Источник
Передвижение воды в растениях

Различные органы растения выполняют разные функции. Листья синтезируют органические вещества, поэтому в них должны постоянно поступать вода и минеральные вещества. Корни поглощают воду и минеральные вещества и нуждаются в притоке органических веществ для дыхания и роста. Цветки, плоды и растущие верхушки растений являются потребителями органических веществ, значительная часть которых откладывается в запас. Все это вызывает необходимость передвижения в растении воды и растворенных в ней веществ.
Вода и растворенные в ней вещества передвигаются в растении в основном двумя путями: путем диффузии и в виде потока. Диффузия воды и веществ осуществляется по градиенту концентрации и подчиняется закону Фика. Движение в виде потока происходит по градиенту гидростатического давления, по градиенту потока вода движется и через мембраны при наличии градиента осмотического или тургорного давления.
Такое передвижение наиболее ярко выражено у древесных растений. Исследования показали, что по стволу дерева вещества передвигаются в двух основных направлениях: от корней к листьям вверх движутся вода и минеральные вещества — восходящий поток; второй, несущий органические вещества вниз к корню, — нисходящий поток. Но органические вещества из листьев поступают не только в корневую систему, они движутся также к морфологической верхушке, цветкам и плодам. Поэтому нисходящий поток лучше назвать потоком пластических веществ.
В существовании двух потоков — восходящего и пластических веществ — можно убедиться на опыте, сущность которого заключается в следующем. Делают кольцевой надрез на стволе дерева или на одном из побегов первого или второго порядка. На побеге вырезают паренхиму коры кольцом шириной в несколько сантиметров. Чтобы избежать высыхания, место выреза обматывают тканью или замазывают садовым варом. Через некоторое время над кольцом вследствие приостановки нисходящего потока образуется наплыв — каллус. Если кольцо не очень широкое, оно обычно срастается.
При образовании такого кольца растение некоторое время хорошо растет, тургесцентное состояние клеток остается нормальным, происходит передвижение воды и растворенных в ней веществ. Наплыв над кольцом образуется вследствие разрастания клеток коровой паренхимы и скопления в них пластических веществ — углеводов, органических кислот и др. Если кольцо достаточно широкое и не срастается, то пластические вещества не будут поступать в корневую систему, она истощится и дерево или побег погибнет.
Восходящий поток идет по сосудам и трахеидам, которые являются мертвыми полыми клетками и сами по себе не обладают сосущей или какой-либо иной силой, способной привести воду в движение. Вследствие вогнутости менисков в сосудах, представляющих собой капилляры, вода может подняться при их диаметре 0,1 мм не выше 30 см. Однако вода в древесных растениях поднимается на десятки метров, поэтому капиллярными силами, транспирацией и корневым давлением это объяснить нельзя.
Вода в сосудах как бы подвешена к испаряющим клеткам в виде тонких нитей. Нижним концом они упираются в сосудах в клетки корневых волосков. Непрерывность водных нитей обусловливается силами взаимного сцепления молекул воды и силами прилипания их к клеточным стенкам сосудов. О существовании водных нитей в растении свидетельствуют многочисленные факты и наблюдения, например уменьшение диаметра стеблей растений и стволов деревьев в результате интенсивной транспирации. Чтобы вода передвигалась вверх, испаряющие клетки должны иметь достаточную сосущую силу, которая в клетках листовой паренхимы бывает довольно большой (достигает 2-4 тыс. кПа и более). Одним из факторов, поддерживающих сосущую силу на высоком уровне, является непрерывная транспирация. Таким образом, движение воды по сосудам объясняется наличием в растении водных нитей, присасывающей силой транспирации и корневым давлением.
Сила сцепления молекул воды в растениях велика. Так, в клетках спорангиев папоротника она превышает 40 тыс. кПа. Исследованиями установлено, что этого вполне достаточно, чтобы не разорвались водные нити, которые заполняют сосудистые полости высокого дерева.
Прочное сцепление (когезия) между молекулами воды и прилипание (адгезия) их к гидрофильным стенкам клеток ксилемы предотвращает образование полостей (кавитацию) в находящемся в ксилеме растворе почти в любых условиях. Однако при сильном дефиците воды в отдельных трубках ксилемы кавитация все же происходит. Убедиться в этом можно следующим образом: если к стволу дерева прижать чувствительный микрофон, то будет слышно потрескивание. Такие трубки ксилемы уже не восстанавливаются, но камбий может образовывать новые.
Силы сцепления молекул воды и присасывающее действие транспирации можно продемонстрировать на таком опыте. Гипсовый блок или ветку сосны герметично присоединяют каучуковой трубкой к пипетке, заполненной водой и погруженной в ртуть. Вода, испаряясь с поверхности гипсового блока или хвои, благодаря силам сцепления между молекулами ртути и воды тянет за собой ртуть, которая и будет подниматься по пипетке. Таким образом, присасывающее действие транспирации и силы сцепления воды в растении обусловливают движение ее на несколько десятков метров.
Вопрос об участии живых клеток древесины в движении воды в растении и связанные с этим явления изучены еще недостаточно. На основании ряда работ считали, что живые клетки древесины и древесной паренхимы имеют способность проталкивать через себя воду, засасывать ее, например, нижним концом и выделять верхним в сосуды, т. е. при этом как бы происходит пульсация воды в клетках. Предполагали, что такое передвижение воды с участием живых клеток идет в заболони— внешнем слое древесины, который прилегает к камбию. Однако дальнейшие исследования этого не подтвердили. Кроме того, считали, что существуют особые клапаны, которые также способствуют перемещению воды в растении. Исследования Е. Ф. Вотчала, изучавшего передвижение воды по стволу дерева, находившегося в горизонтальном положении, этого не подтвердили.
Некоторые исследователи (Т. Беннет-Кларк, Д. Биксон, Р. Гёбер и др.) важным фактором в регулировании водного баланса клеток считают электроосмос. Это — движение молекул воды, несущих электрический заряд вдоль поверхности раздела (например, по стенкам пор мембран). Электроосмотическое движение жидкости происходит в результате возникновения двойного. электрического слоя на поверхности раздела жидкость—твердое тело. При погружении мембраны в раствор, содержащий электролиты, ее поверхность приобретает заряд (для естественных мембран обычно отрицательный), который возникает в результате процесса ионизации поверхности мембран или вследствие адсорбции ионов из раствора. Жидкость возле поверхности мембраны несет заряд, противоположный по знаку, и образует слой, обладающий подвижностью. Он обусловлен разностью потенциалов, вызванной наличием в растворе электролитов, соприкасающихся с мембраной.
Аномальное движение частиц может приводить к превышению наблюдаемого давления над осмотическим, обусловленному электроосмотическим потоком воды через поры мембраны. Для осуществления электроосмотического потока жидкости необходимо наличие проницаемой мембраны, имеющей систему пор различной величины; электролитов в соответствующих концентрациях по обе стороны мембраны; постоянной диффузии электролитов.
Абсолютная скорость передвижения воды по древесине у лиственных пород составляет 20 см 3 , хвойных — 5 см 3 на 1 см 2 поперечного среза древесины в час. В опытах с мечеными атомами установлено, что скорость движения воды по ксилеме составляет 12-14 м/ч.
При перемещении воды по дереву по вертикали в гравитационном поле водный потенциал возрастает примерно на 1 бар до 10 м, так как известно, что гидростатическое давление 101,3 кПа может поддерживать столб ртути высотой 76 см или столб воды 10,3 м. Но вода в деревьях поднимается на 20-40 м и выше, что обусловливается преимущественно величиной сосущей силы транспирации и силами сцепления молекул воды.
Наблюдения показали, что в жаркое время дня имеющиеся в листьях запасы воды полностью обновляются приблизительно каждый час. Такой высокий уровень расхода влаги растением может быть обеспечен лишь при большой скорости движения ее по ткани.
Таким образом, благодаря верхнему (транспирация) и нижнему (корневое давление) двигателям водного потока и силам сцепления молекул в сосудах происходит передвижение и поднятие воды по растению на большую высоту. Путь, который проходит вода по растению, делится на две неравные части: первая — движение воды по сосудам и трахеидам (этот путь составляет от нескольких сантиметров до нескольких метров); вторая — движение водного потока по живым клеткам (его протяженность выражается в миллиметрах и даже долях миллиметра). Во вторую часть водного потока входят два коротких участка: первый — в корне, от корневого волоска до сосуда, находящегося в центральном цилиндре; второй — в листе, от жилок до испаряющих клеток мезофилла (рис. 20).

Рис. 20. Начальный и конечный отрезки пути водного потока в растении:
A — корень; Б — лист (простыми стрелками показан путь, который проходит вода, оперенными — путь водяного пара).
Движение воды по сосудам можно показать на таком примере. Букет цветов сохраняет свежесть, если вода будет свободно поступать в перерезанные сосуды. Поэтому, чтобы в стеблях не образовывались так называемые воздушные пробки, рекомендуется обрезать их на 5-10 см под водой.
По сосудам вода течет, как по полым трубкам, подчиняясь общим законам гидродинамики. В паренхимных клетках вода движется осмотическим путем, и передвижение ее в живых клетках значительно затруднено. Но основным двигателем водного потока в растениях является сосущая сила паренхимных клеток листьев, или присасывающее действие транспирации.
О природе приспособительных реакций к недостатку воды у разных групп растений. Транспирирующие органы — листья — характеризуются значительной пластичностью, в зависимости от условий произрастания в их строении наблюдаются довольно большие изменения. Даже листья одного растения при разных условиях водоснабжения и освещения имеют различия в анатомической структуре.
Установлены определенные закономерности в строении листьев в зависимости от расположения их на растении. В. Р. Заленский обнаружил изменения в анатомическом строении листьев по ярусам. Он установил, что у листьев верхнего яруса наблюдаются закономерные изменения в сторону усиления ксероморфизма, т. е. образуются структуры, повышающие засухоустойчивость этих листьев. Установленные им закономерности называют законом Заленского. Листья, расположенные в верхней части стебля, всегда отличаются от нижних, а именно: чем выше расположен лист на стебле, тем меньше размеры его клеток, тем больше на нем устьиц и меньше их размеры, больше волосков на единицу поверхности, гуще сеть проводящих пучков, сильнее развита палисадная ткань. Все эти признаки характеризуют ксерофилию, т. е. образование структур, способствующих повышению засухоустойчивости.
С определенной анатомической структурой связаны и физиологические особенности, а именно: верхние листья отличаются более высокой ассимиляционной способностью и более интенсивной транспирацией. Концентрация сока в верхних листьях также более высокая, в связи с чем может происходить оттягивание воды верхними листьями от нижних, засыхание и отмирание нижних листьев. Структура органов и тканей, обусловливающая повышение засухоустойчивости растений, называется ксероморфизмом. Отличительные особенности структуры листьев верхнего яруса объясняются тем, что они развиваются в условиях несколько затрудненного водоснабжения.
Физиологические причины ксероморфной структуры следует рассматривать как изменения цепи ферментативных реакций, вызванных недостатком воды и приводящих к торможению роста клеток в фазе растяжения.
Ксероморфная структура листьев растений вызывается тем, что дефицит воды проявляется прежде всего в период ранней приостановки роста эпидермиса — эпидермальных клеток. В нормальных условиях фаза растяжения приостанавливается вначале на нижнем мезофилле.
Для уравнения баланса между поступлением и расходом воды в растении образовалась сложная система анатомо-физиологических приспособлений. Такие приспособления наблюдаются у ксерофитов, гигрофитов, мезофитов. Большой интерес в связи с этим представляют исследования Б. А. Келлера, который изучал анатомо-физиологические особенности у растений резко различающихся экологических групп, но близких между собой в систематическом отношении. Он исследовал многолетние травянистые растения (род Asperula) из семейства Мареновые. Одни из них были типичными степными растениями, а другие — теневыносливыми, лесными.
Степной вид ясменника колокольчиковидного (Asperula glauca) имеет сизые узкие иглообразные толстые листья с сильно развитой двуслойной палисадной паренхимой, края листьев немного загнуты вниз. Это типичный ксерофит, растущий на открытой местности в степной зоне или полупустынях. Второй вид — ясменник душистый (Asperula odorata) — характерен для влажных и очень тенистых участков лиственного леса; у растений этого вида широкая и тонкая пластинка, однослойная палисадная ткань, состоящая из укороченных и рыхлорасположенных клеток. Было проведено сравнительное изучение анатомического строения и интенсивности транспирации у обоих видов (табл. 4).
| Вид | Длина сети жилок листа, мм | Количество устьиц | Интенсивность транспирации, % |
| Asperula glauca | 100 | 100 | 100 |
| Asperula odorata | 30 | 14 | 45 |
Такие же исследования провел Б. А. Келлер с двумя видами подмаренника: весенним (Galium verum) и крестовидным (Galium cruciata). Полученные им данные также свидетельствуют о том, что условия произрастания оказывают большое влияние на анатомическую структуру и физиологические особенности растения. Если ксерофиты одновременно являются гелиофитами, то мезофиты и даже гигрофиты необязательно относятся к теневыносливым растениям.
При выращивании фасоли (опыт Н. А. Максимова) на различном расстоянии от источника света (электрическая лампа) было установлено, что у сильнее освещенных экземпляров устьиц на листе было приблизительно в 4 раза больше, чем у плохо освещенных, размеры клеток эпидермиса в 3-4 раза меньше и сеть жилок значительно гуще. Таким образом, степень освещенности и нагревания сильно влияет на анатомическое строение.
Н. А. Максимов проведенными исследованиями опроверг господствовавшие в физиологии того времени взгляды на засухоустойчивость как на биологически обоснованную потребность растения в недостаточном водоснабжении, как на сухолюбне. Его исследования показали, что засухоустойчивость следует понимать как приспособительное свойство растений переносить глубокое завядание с наименьшим вредом не только для данной особи, но и для всего вида.
Различие между стойкими и нестойкими к засухе растениями обусловлено характером тех изменений в обмене веществ, которые возникают у растения под влиянием обезвоживания. Так, уровень обводненности ткани у неустойчивых к засухе сортов пшеницы выше по сравнению с устойчивыми. Кроме того, у неустойчивых сортов наблюдается более высокий уровень гидролитического действия ферментов углеводного и белкового обменов. Однако эти признаки непостоянны и проявляются лишь в условиях полного насыщения водой, а при нарушении водоснабжения быстро утрачиваются. Результаты исследований показали, что приспособительные свойства у засухоустойчивых форм растений возникают под влиянием условий их существования.
Источник