Поддержание водного баланса растениями
Низшие наземные растения из влажного субстрата поглощают воду погруженными в него частями таллома, а влагу дождя, росы и тумана – всей поверхностью. В максимально набухшем состоянии лишайники содержат в 20–30 раз больше воды, чем сухого вещества.
Среди высших наземных растений мохообразные поглощают воду из почвы ризоидами, а большинство других – корнями, специализированными органами, всасывающими воду. В клетках корня развивается сосущая сила чаще всего в несколько атмосфер , но этого достаточно для извлечения из почвы большей части связанной воды. Лесные деревья умеренной зоны развивают сосущую силу корней около 3 · 10 6 Па (30 атм ), некоторые травянистые растения (земляника лесная, медуница неясная) – до 2 · 10 6 (20 атм ) и даже свыше 4 · 10 6 Па (40 атм ) (смолка обыкновенная); растения сухих областей – до 60 атм.
Когда в непосредственной близости от корней запасы воды в почве истощаются, корни растут в направлении большей влажности, так что корневая система растений постоянно находится в движении. У степных и пустынных растений часто можно видеть эфемерные корни, быстро вырастающие в периоды увлажнения почвы, а с наступлением засушливого периода засыхающие.
По типу ветвления различают следующие корневые системы:
1) экстенсивная охватывает большой объем почвы, но сравнительно слабо ветвится, так что почва пронизана корнями негусто. Таковы корневые системы у многих степных и пустынных растений (саксаула, верблюжьей колючки), у деревьев умеренной полосы (сосны обыкновенной, березы повислой ), а из трав у люцерны серповидной, василька шероховатого и др.;
2) интенсивная– ох ватывает сравнительно небольшой объем почвы, но густо пронизывает ее многочисленными сильно ветвящимися корнями, как, например, у степных дерновинных злаков (ковылей, типчака и др.), у ржи, пшеницы. Между этими типами корневых систем есть переходные .
Корневые системы очень пластичны и резко реагируют на изменение условий, в первую очередь увлажнения. При недостатке влаги корневая система становится экстенсивнее . Так, при выращивании ржи в разных условиях общая длина корней (без корневых волосков) в 1000 см 3 почвы варьирует от 90 м до 13 км , а поверхность корневых волосков может увеличиться в 400 раз.

Рис. Корневые системы стенных и тундровых растений:
А – Festuca sulcata ; Б – Euphorbia gerardiana на черноземах в Аскания-Нова ;
В – Eriophorum scheuchzeri ; Г – Hierochloe alpina – из тундр Таймыра
Всасывание воды корнями затруднено при большой сухости почвы, засолении или сильной кислотности, при низкой температуре. Например, ясень обыкновенный при температуре почвы 0 °C поглощает воды в 3 раза меньше, чем при +(20–30) °С. Способность поглощать воду при той или иной температуре зависит от приспособленности растений к тепловому режиму почв в местах их произрастания. Виды с ранним началом развития, как правило, могут всасывать воду корнями при более низкой температуре, чем развивающиеся позднее. Тундровые растения и некоторые деревья, растущие на почвах с подстилающей их многолетней мерзлотой, могут поглощать воду при температуре почвы 0 °C .
У высших растений есть и дополнительные пути поступления воды в тело. Мхи могут поглощать воду всей поверхностью, как и лишайники. Особенно много воды впитывают такие мхи, как кукушкин лен, виды сфагнума, чему способствует строение их листьев и побегов. При полном насыщении сфагновые мхи содержат в своем теле в десятки раз больше воды, чем в воздушно-сухом состоянии.

Рис. Часть листа сфагнума болотного с клетками двух типов: зеленых с хлоропластами и бесцветных водоносных
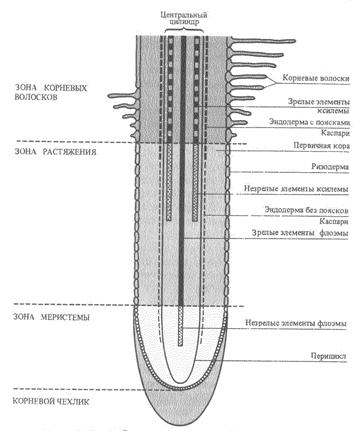
Семена поглощают воду из почвы. Из воздуха, насыщенного водяными парами, в дождевом тропическом лесу поглощают воду многие эпифиты, например папоротник гименофиллум – тонкими листьями, многие орхидеи – воздушными корнями. В чашевидных влагалищах листьев многих зонтичных скапливается вода, которая постепенно всасывается эпидермисом. Виды из рода тилляндсия ( бромелиевые ) существуют в пустыне Атакама практически исключительно за счет влаги туманов и росы, которую впитывают чешуевидные волоски на листьях.
Поступившая в растение вода транспортируется от клетки к клетке (ближний транспорт) и по ксилеме во все органы, где расходуется на жизненные процессы (дальний транспорт). В среднем 0,5 % воды идет на фотосинтез, а остальная – на восполнение потерь от испарения и поддержание тургора. Вода испаряется со всех поверхностей, как внутренних, так и наружных, соприкасающихся с воздухом. Различают устьичную , кутикулярную и перидермальную транспирацию.
Через устьица транспирируется влага, испарившаяся с поверхности клеток внутри органов. Это основной путь расходования воды растением. Кутикулярная транспирация составляет менее 10 % от свободного испарения; у вечнозеленых хвойных пород она сокращается до 0,5 %, а у кактусов даже до 0,05 %. Относительно велика кутикулярная транспирация молодых развертывающихся листьев. Перидермальная транспирация обычно незначительна. Интенсивность общей транспирации повышается с увеличением освещенности, температуры, сухости воздуха и при ветре.
Водный баланс остается уравновешенным в том случае, если поглощение воды, ее проведение и расходование гармонично согласованы друг с другом. Нарушения его могут быть кратковременными или длительными. По приспособлениям наземных растений к кратковременным колебаниям условий водоснабжения и испарения различают пойкилогидрические и гомойогидрические виды.
У пойкилогидрических растений содержание воды в тканях непостоянно и сильно зависит от степени увлажнения окружающей среды. Они не могут регулировать транспирацию и легко и быстро теряют и поглощают воду, используя влагу росы, туманов, кратковременных дождей, в сухом состоянии находятся в анабиозе. Способны обитать там, где короткие периоды увлажнения чередуются с длительными периодами сухости.
Пойкилогидричность свойственна цианобактериям , всем водорослям, некоторым грибам, лишайникам, а также ряду высших растений: многим мхам, некоторым папоротникам и даже отдельным цветковым, по-видимому, вторично перешедшим к пойкилогидрическому образу жизни. Таков, например, южно-африканский кустарник Myrothamnus flabel-lifolia ( розоцветные ).

Рис. Лишайник сосновых лесов Cetraria islandica – типичный пойкилогидрический организм
В мелких клетках таллома большинства низших растений нет центральной вакуоли, поэтому при высыхании они равномерно сжимаются без необратимых изменений ультраструктуры протопласта. Сине-зеленые водоросли ( цианобактерии ), вегетирующие на поверхности почвы в пустыне, высыхая, превращаются в темную корочку. От редких дождей их слизистая масса набухает и нитчатые тела начинают вегетировать . Мхи, растущие на сухих скалах, стволах деревьев или на поверхности почвы лугов и степей (роды Thuidium , Tortula и др.), также могут сильно высыхать, не теряя жизнеспособности.
Пойкилогидричны пыльцевые зерна и зародыши в семенах растений.
Гомойогидрические растения способны поддерживать относительное постоянство обводненности тканей. К ним относят большинство высших наземных растений. Для них характерна крупная центральная вакуоль в клетках. Благодаря этому клетка всегда имеет запас воды и не так сильно зависит от изменчивых внешних условий. Кроме того, побеги покрыты с поверхности эпидермой с малопроницаемой для воды кутикулой, транспирация регулируется устьичным аппаратом, а хорошо развитая корневая система во время вегетации может непрерывно поглощать влагу из почвы. Однако способности растений, не выдерживающих высыхания, регулировать свой водный обмен различны . Среди них выделяют разные по экологии группы.
Источник
Поглощение воды растением




![]()
![]()
Корневая система является главным аппаратом поглощения воды, но не единственным: небольшое количество воды поглощается надземными частями растения во время дождей, выпадения туманов, росы. В принципе любая клетка может всасывать воду, если она не насыщена ею, даже кутикула способна разбухать.
Размеры корневых систем растений огромны. Хлебные злаки способны развивать корневую систему, достигающую глубины 1,5 — 2 м и широко распространяющуюся в стороны. Глубина залегания корней древесных растений может достигать 20 м и более (песчаная акация, тамарикс и др.). у столетней сосны насчитывается около 5 млн корневых окончаний, а общая длина корневой системы (без корневых волосков) достигает 50 км.
Все многообразие древесных растений по характеру строения и развития корневых систем делят на три основные группы:
· с глубоко идущей в почву якорной корневой системой (лиственница, дуб, липа, робиния и др.);
· с поверхностной корневой системой (ель, ясень, рябина и др.);
· с промежуточным типом корневых систем, включающим в зависимости от лесорастительных условий как глубоко идущую, так и поверхностную корневую систему (сосна, береза, осина, многие клены, ольха и др.).
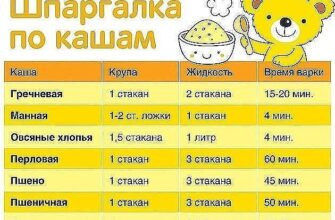
Далеко не вся корневая система всасывает воду. Этой способностью обладают лишь молодые корневые окончания. Вода поглощается клетками зоны корневых волосков, а также зоны растяжения. Поглощающие корни относятся к последнему порядку ветвления. Для них характерно первичное строение, отсутствие камбиального слоя. Основная их функция поглощение воды и элементов минерального питания из почвы, тогда как для ростовых — увеличение размеров корневых систем.
Длина всасывающей части корня травянистых растений достигает нескольких сантиметров, а древесных — всего лишь нескольких миллиметров. При этом у большинства деревьев всасывающий аппарат увеличивается за счет развития микоризы — симбиоза корня с гифами гриба. Грибной чехол микоризных корней и отходящие от него отдельные гифы существенно увеличивают поглощающую поверхность корня. Оказалось также, что микоризные растения более устойчивы к засухе, чем растения без микоризы: грибница продолжает активно всасывать воду при таком осмотическом давлении почвенного раствора, когда безмикоризные корни уже не работают.
Различают пассивное и активное поглощение воды корневой системой. Первое осуществляется за счет сосущих сил листьев, теряющих воду в процессе транспирации. Активное поглощение тесно связано с метаболизмом клеток корня и проявляется в явлениях «плача» и гуттации.
При перерезании стебля через поверхность среза начинает вытекать жидкость — пасока. Это явление носит название «плача» растений. Очень легко наблюдается «плач» у таких растений, как тыква, гортензия, а из древесных — у березы, кленов, особенно сахарного.
По многолетним данным В.П. Рябчука, сезонная сокопродуктивность одного дерева березы в различных районах страны (в зависимости от диаметра и количества подсочных каналов) составляет 37 — 360 л.
В пасоке березы при весеннем сокодвижении содержится до 1,5 % сахаров, а у отдельных деревьев до 3,5 % (сахароза, глюкоза и фруктоза). В небольших количествах в березовом соке обнаруживаются аминокислоты, жирные кислоты, минеральные соли и некоторые другие вещества. Эти качества пасоки широко используют в народном хозяйстве. Она добывается в процесс е так называемой ксилемной подсочки берез, кленов и некоторых других пород.
Под ксилемной подсочкой понимается совокупность производственных процессов, связанных с добычей ксилемного сока лиственных древесных растений. Добываемая жидкость (пасока) используется для приготовления сока, сиропа, кваса, вина, спирта, а также в качестве добавок в парфюмерно-косметическом производстве, медицине, животноводстве и пчеловодстве. Более
всего заготавливалось березового сока среди стран СНГ на Украине (в среднем 35 тыс. т/год), Белоруссии и России. Грамотное ведение ксилемной подсочки березы в 5 раз экологически более эффективно, чем сбыт березовой древесины с единицы лесопокрытой площади. Еще более выгодна пневая подсочка берез. Ксилемная подсочка лиственных древесных растений существенно снижает и затраты картофеля и других ценных с/х продуктов в спиртовом производстве.

Иногда можно наблюдать выделение капелек жидкости из неповрежденных листьев. Это гуттация. Она ярко проявляется у проростков злаков, У каллы, манжетки, земляники. Из древесных растений нередко гуттируют ольха, ивы, вязы и некоторые другие. Гуттация обусловлена тем, что при высокой влажности воздуха и почвы в теплую погоду вода, поступившая в растение, не испаряется, а выдавливается в виде капелек жидкости. У срезанного побега дерева или кустарника даже при 100 % влажности воздуха гуттация наблюдаться не будет, ибо отсутствует корневое давление.
«Плач» и гуттация связаны с наличием корневого давления, нагнетающего воду вверх из корней по растению. Само корневое давление является результатом жизнедеятельности корня. Движущей силой корневого давления служит энергия аэробного дыхания, ибо ингибиторы дыхания (яды, наркотики) и анаэробиоз приводят к прекращению корневого давления.
Роль процесса дыхания в проявлении корневого давления может быть не только прямой, но и косвенной. Энергия этого процесса способствует поглощению ионов солей из почвы, передвижению их по живым клеткам корня и выделению в полости сосудов центрального цилиндра корня. При интенсивном дыхании концентрация солей в сосудах корня увеличивается, а вместе с ней повышается осмотическое давление раствора в сосудах.
Существуют две основные гипотезы, объясняющие механизм действия корневого давления:
Осмотuческая теория основана на явлениях осмоса. В сосудах центрального цилиндра поглощающего корня всегда находится раствор определенной концентрации, обладающий сосущей силой, равной целиком осмотическому давлению, так как в мертвых элементах ксилемы отсутствует тургорное давление. Поэтому вода будет отсасываться от ближайшей к сосуду клетки. Теряя воду, эта клетка будет развивать большую сосущую силу, что позволит ей отнимать воду от соседней с ней клетки и т.д. Клетки коровой паренхимы и центрального цилиндра корня будут испытывать недонасыщение водой. Этот дефицит коснется и корневого волоска, способного поглощать воду из почвы. При этом распределение величины осмотического давления в ряду клеток, про водящих воду, не играет никакой роли, ибо основным фактором осмотического поглощения воды выступает сосущая сила клеток.
Метаболuческая теория, выдвинутая Д.А Сабининым, основывается на допущении, что клетки корня, через которые наблюдается односторонний ток воды, обладают полярностью. На противоположных сторонах осуществляются диаметрально разные процессы: на одной (обращенной к периферии) синтезируются осмотически активные сахара, а на противоположной — инертный в осмотическом отношении крахмал. В первом случае осмотическое давление и сосущая сила будут больше, следовательно, к этой стороне клетки будет поступать вода от соседней клетки и выдавливаться с противоположного конца в сторону сосуда. Так создается непрерывный односторонний ток воды в корневой системе, как результат развиваемого в корнях корневого давления.
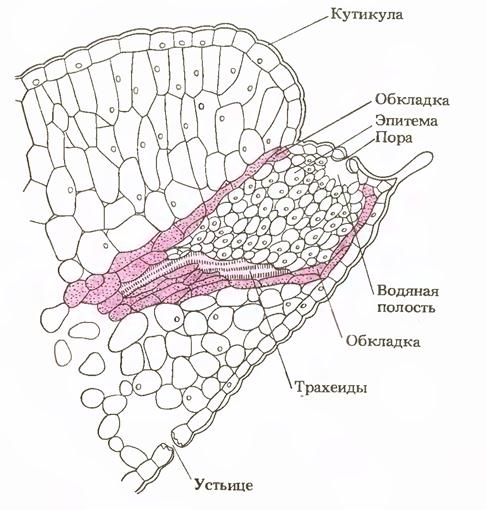
В области эндодермы корня вода проходит через ее пропускные клетки, но их сравнительно мало. Другие клетки эндодермы с суберинизированными поясками Каспари, тесно связанными с плазмалеммой, представляют сильное препятствие току воды и растворов. По существу, эндодерма — основной барьер, препятствующий одностороннему току воды из клеток первичной коры в центральный цилиндр корня. Недавно с помощью современной техники было показано, что растворы проходят только через протопласты клеток эндодермы, тогда как в других клетках — как через симпласт, так и через апопласт (по клеточным стенкам).
Величина корневого давления обычно колеблется около 1 атм, реже 2,5 — 3,0 атм. С помощью корневого давления растения в ночное время суток восполняют водный дефицит, возникающий в жаркий летний день.
Корневое давление, у древесных растений действует в течение всего года, в том числе зимой, пополняя водные запасы ствола. Как установил Л. А. Иванов, после спиливания дерева влажность древесины пня повышается под действием корневого давления. Особенно велико значение корневого давления ранней весной до распускания листьев, когда отсутствует притягивающая воду транспирация. Корневое давление можно определить с помощью манометра, прикрепленного к пеньку срезанного растения.
В обычных условиях роль пассивного поглощения воды в водообмене растений существенно выше. Оно более выгодно и в энергетическом отношении, так как не связано с затратой внутренней энергии, присущей активному поглощению, а осуществляется за счет энергии Солнца.
Источник