Основные закономерности поглощения воды клеткой.
Растительная клетка окружена эластичной клеточной стенкой. Вакуоль содержит большое количество осмотически активных веществ. При изучении осмотических явлений в растительных клетках полупроницаемой мембраной считается система, состоящая из плазмалеммы и тонопласта одновременно. При помещении клетки в воду последняя по законам осмоса будет поступать внутрь клетки.
Силу, с которой вода входит в клетку, называют сосущей силой S. Она тождественна водному потенциалу клетки (Ψв). Величина сосущей силы определяется осмотическим давлением клеточного сока (π*) и тургорным (гидростатическим) давлением в клетке (Р), которое равно противодавлению клеточной стенки, возникающему при ее эластическом растяжении:
Когда клетка полностью насыщена водой (полностью тургесцентна), ее сосущая сила равна нулю, а тургорное давление равно потенциальному осмотическому: S = 0; Р = π*. Если подача воды к клетке уменьшается (при усилении ветра, при недостатке влаги в почве и т. д.), то вначале возникает водный дефицит в клеточных стенках, водный потенциал которых становится ниже, чем в вакуолях, и вода начинает перемедцаться в клеточные стенки. Отток воды из вакуоли снижает тургорное давление в клетках и, следовательно, увеличивает их сосущую силу. При длительном недостатке влаги большинство клеток теряет тургор и растение подвядает. В этих условиях Р = 0; S=π*.

Схема растительной клетки как осмотической системы:
π* — осмотическое давление, Р — тургорное давление, —Р — противодавление клеточной стенки
Явление потери тургора клетками можно наблюдать, если поместить кусочки тканей в гипертонический раствор. Отток воды из клеток приводит к уменьшению объема протопластов и их отделению от клеточных стенок. Происходит плазмолиз. Пространство между стенкой и сократившимся протопластом заполняет наружный раствор. При помещении этой клетки в чистую воду тургересцентность клетки восстанавливается.
В условиях водного дефицита в молодых тканях резкое усиление потери воды, например при суховее, может приводить к тому, что тургорное давление в клетках становится отрицательной величиной и протопласты, сокращаясь в объеме, не отделяются от клеточных стенок, а тянут их вслед за собой. Клетки и ткани сжимаются. Это явление называется циторризом.
Источник
Поглощение воды клеткой
Поглощение питательных веществ клеткой.
Для осуществления всех процессов жизнедеятельности в клет-ку из внешней среды должны поступать вода и питательные веще-ства. Цитоплазма и ее пограничные мембраны — плазмалемма и тонопласт — обладают свойствами избирательной проницаемости. Под избирательной проницаемостью понимают способность актив-‘1ю регулировать процесс поступления веществ: одни вещества пропускать с большей скоростью, другие — с меньшей или не пропускать совсем. Таким путем биологическая система может сохранять стабильность — гомеостаз и не смешиваться с окружа-ющей средой.
Поглощение может быть пассивным и активным. Пассивным называется перемещение веществ путем диффузии по электрохи-мическому (концентрационному и электрическому) градиенту.
Диффузия — движение молекул или ионов из области с высокой концентрацией в область с более низкой концентрацией. Ионы перемещаются через мембраны в том или ином направлении бла-годаря своей собственной кинетической энергии, не потребляя в этом процессе ни АТФ, ни другие источники энергии. Некоторые молекулы проникают растворяясь в липидах мембран или через специальные гидрофильные канальцы в мембранах. Так как ионы заряжены, скорость их диффузии определяется не только прони-цаемостью мембраны и различиями в концентрации ионов по обе ее стороны, но также электрическим потенциалом — напряжени-ем, возникающим между внутренней и внешней сторонами мем-браны. Обычно растительные клетки на внутренней стороне своих мембран имеют отрицательный потенциал, что приводит к пре-имущественному поглощению положительно заряженных ионов по сравнению с отрицательными.
Активный транспорт — это поглощение веществ через специ-фические зоны мембраны, называемые насосами, против элект-рохимического градиента с затратой энергии дыхания. Например, растительная клетка поглощает калий в количествах в сотни и тысячи раз больших, чем он имеется в окружающей среде.
Вода может поступать в клетки растений благодаря набуханию биоколлоидов, увеличению степени их гидратации. Это характер-но для сухих семян, помещенных в воду. Однако главный способ поступления воды в живые клетки — ее осмотическое поглоще-ние.
Большая часть присутствующей в клетке воды находится в ва-куоле. Там же содержится большое количество других веществ— сахаров, органических кислот, солей. Для того чтобы попасть в вакуоль, вода должна пройти сквозь стенку клетки, а также через две мембраны и лежащую между ними цитоплазму. Обычно эти три структуры рассматривают совместно как единый мембранный барьер, обладающий свойствами полупроницаемости, т. е. прони-цаемостью только для воды. Согласно молекулярно-кинетической теории, молекулы всех веществ находятся в состоянии быстрого хаотического движения, скорость которого зависит от энергии этих молекул. Молекулы воды малы и проходят через клеточные мемб-раны намного быстрее, чем молекулы других веществ. Они диф-фундируют во всех направлениях: в клетку и из клетки, в различ-ные клеточные органеллы и из них. Движение воды осуществляет-ся по градиенту ее активности„ которая характеризуется водным потенциалом. Молекулы содержащихся в вакуоле растворенных веществ снижают активность молекул воды в клетке. Вследствие этого кинетическая энергия воды в вакуоле ниже, чем кинетичес-кая энергия относительно более чистой воды вне клетки. В резуль-тате внутрь клетки в вакуоль проникает больше молекул воды, чем выходит из нее. Объем вакуоли увеличивается и создается тургор, в результате которого содержимое клетки прижимается к ее стенке.
Диффузия воды через полупроницаемую мембрану называется осмосом. Если два раствора разделены полупроницаемой мембра-ной, то вода из раствора с меньшей концентрацией веществ (ги-потонического) будет диффундировать в раствор с большей кон-центрацией веществ (гипертонический). Диффузия будет проис-ходить до тех пор, пока концентрации веществ в обоих растворах не станут одинаковыми (изотоническими).
Если в осмометре раствор сахарозы отделить от чистой воды полупроницаемой мембраной (рис. 8.2), то молекулы воды в силу диффузии будут перемещаться из чистой воды в раствор сахаро-зы, где концентрация воды меньше, т.е. от высокого водного по-тенциала к более низкому. Это приводит к увеличению объема раствора сахарозы и подъему жидкости в трубке осмометра. При этом повышается гидростатическое давление, под которым нахо-дится раствор в осмометре. При некоторой высоте столба раствора в трубке скорости диффузии воды сравняются и подъем его пре-кратится.
Наибольшее давление, которое способен преодолеть данный раствор, всасывая воду через полупроницаемую мембрану, яв-ляется его осмотическим давлением. Осмотический потенциал ра-створа прямо зависит от концентрации растворенного вещества и по величине равен потенциалу гидростатического давления, по противоположен ему по знаку. Он всегда имеет отрицатель-ное значение. Величина осмотического потенциала неоднознач-на у разных видов растений и в разных частях одного и того же растения. Для вакуолярного сока клеток корней она составляет 12 -13 ат., а для клеток надземных органов 10 – 26 ат. Это обуславливает вертикальный градиент асмотического потенциала и сосущей силы от корней к листьям. У растений- галофитов, растущих в условиях засоления, осмотический потенциал очень высок, достигая 150 ат.
Силу, с которой вода входит в клетку, называют сосуищей силой (5), она тождественна водному потенциалу клетки. Величина со-сущей силы определяется осмотическим давлением клеточного сока (z) и тургорным (гидростатическим) давлением в клетке
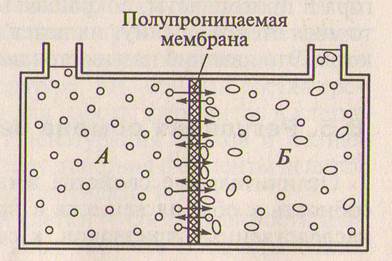
Рис. 8.2. Схема простейшего осмо-метра:
1 — сосуд с водой; Б — сосуд с 10%-м
раствором сахарозы (кружки — молеку-лы воды, овалы — молекулы сахарозы).
(Р), которое равно противодавлению клеточной стенки, возникающе-му при ее эластичном растяжении: S = к — Р.
В условиях разной оводненности соотношение между всеми компонентами этого уравнения меняется. Когда клетка полнос-тью насыщена водой (полностью тургесцентна), ее сосущая сила равна нулю, а тургорное давление равно потенциальному осмо-тическому: S= 0; Р= к. Состояние полного тургора наблюдается в клетках водных растений. Клетки наземных растений из-за транс-пирации почти никогда не бывают полностью насыщенными во-дой. Если подача воды к клетке уменьшается (при усилении вет-ра, при недостатке влаги в почве и т.д.), то вначале возникает водный дефицит в клеточных стенках, водный потенциал кото-рых становится ниже, чем в вакуолях, и вода начинает переме-щаться в клеточные стенки. Отток воды из вакуоли снижает тур-горное давление в клетках и, следовательно, увеличивает их сосу-щую силу. При длительном недостатке влаги большинство клеток теряет тургор и растение завядает. В этих условиях Р = 0; S = к. Обезвоживание клетки называется плазмолизом.

Явление плазмолиза можно наблюдать, помещая кусочки тка-ни в гипертонический раствор. Отток воды из клеток приводит к уменьшению объема протопластов и их отделению от клеточных стенок. Пространство между клеточной стенкой и сократившимся протопластом заполняет наружный раствор, который легко про-ходит через клеточную стенку, практически не препятствующую движению воды. Клетку в таком состоянии называют плазмолизи-рованной. При помещении клетки в чистую воду она может восста-новить свой тургор. Чрезмерный дефицит приводит к разрыву про-топластов и гибели клеток.
В условиях водного дефицита в молодых тканях резкое усиле-ние потери воды, например при суховее, приводит к потере тур-гора и протопласты, сокращаясь в объеме, не отделяются от кле-точных стенок, а тянут их вслед за собой. Клетки и ткани сжима-ются. Это явление называется циторризом.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Источник
Поглощение воды растительной клеткой

Коллоидные и осмотические свойства растительной клетки в значительной мере определяют законы проникновения в нее воды из окружающей среды. Рассмотрим, что происходит с сухим: семенем, если его поместить в воду или во влажную почву. Обычно такие семена набухают, увеличиваются в объеме, поскольку клеточные оболочки, протопласт и запасные вещества представляют высохшие коллоидные студни, или мицеллы, с большим содержанием белковых веществ, которые хорошо поглощают воду. Сильнее набухают белковые вещества, меньше — крахмал, еще слабее — клетчатка. Поэтому сильнее набухают семена, богатые белками, например семена фасоли и гороха при набухании увеличиваются в объеме почти вдвое. Значительно меньше набухают семена злаков.
Сила, с которой воздушно-сухие семена бобовых растений поглощают воду, достигает 1013 кПа и более. Способность сухих семян с огромнейшей силой поглощать воду и отнимать ее от других более влажных тел, например от почвы, имеет большое значение при их прорастании. До начала прорастания семя поглощает воду лишь вследствие набухания имеющихся в нем коллоидов, что приводит к разрыву семенной оболочки. С началом роста корешка и других эмбриональных частей появляются клетки с вакуолями, и далее насасывание воды прорастающим семенем обусловливается не только набуханием коллоидов, но и явлением осмоса. Клеточная оболочка растягивается ограниченно: расширяясь от поступления воды, она проявляет эластичное противодавление на содержимое клетки и уравновешивает ее осмотическое давление. Наступает такой момент, когда противодавление клеточной оболочки уравновешивает гидростатическое давление клеточного сока. Это происходит при полном насыщении клетки водой. Таким образом, благодаря осмотическому притоку воды в вакуоль в клетке возникает гидростатическое давление клеточного сока, называемое тургорным давлением. При полном насыщении клетки водой осмотическое давление (Р) будет равно тургорному (Т):
Р = Т, или Р — Т = 0.
При испарении воды из клеток объем их уменьшается, а осмотическое давление возрастает, поскольку в меньшем объеме содержится такое же количество растворенных в воде веществ. Следовательно, при испарении воды из клеток осмотическое давление выше тургорного, и в этом случае уравнение можно написать так:
P = T + S.
Величина S — давление, которое обусловливает приток воды в вакуоль, и называется сосущей силой (водный потенциал Ψ):
S = P — T.
У растений, находящихся в завядшем состояний, тургор отсутствует и S равно Р, т. е. величина S соответствует величине осмотического давления.
Таким образом, уровень сосущей силы клетки определяет поступление воды в нее. Растительную клетку можно рассматривать как саморегулирующийся осмотический механизм, который всасывает воду тем сильнее, чем больше ее требуется. При значительной потере воды осмотическое давление и сосущая сила возрастают и поступление воды становится более интенсивным. Соотношения между тургором, осмотическим давлением и сосущей силой можно изобразить графически (рис. 12). Для этого на оси абсцисс откладывают степень растяжения клетки, а на оси ординат — осмотическое давление. Если растение находится в увядшем состоянии, то степень растяжения оболочки клетки будет наименьшей и равна единице. При насыщении клетки водой оболочка может растягиваться лишь в 1,5 раза. При наименьшем растяжении клеточной оболочки сосущая сила клетки достигает максимального значения и равна осмотическому давлению, т. е. при потере тургора осмотическое давление наивысшее (оно отвечает величине сосущей силы). Далее при увеличении тургора сосущая сила будет уменьшаться. Итак, с увеличением тургорного давления сосущая сила уменьшается, и при максимальном тургоре, т. е. при полном насыщении клетки водой, она будет равна 0.

Рис. 12. Схема изменения осмотических величин в клетке при переходе от завядания к насыщению водой (слева направо):
Т — тургорное давление; О — осмотическое давление клеточного сока; S — сосущая сила.
Однако в отдельных случаях сосущая сила клетки может быть выше осмотического давления. Так, при увядании растений плазмолиза в клетках не наблюдается. Протопласт таких клеток уменьшается в объеме, но не отделяется от оболочки, а тянет ее за собой. Внешне это проявляется в образовании на поверхности клеток волнообразных изгибов. Такое явление называется циторризом. В клетках увядших растений тургорное давление меньше 0, т. е. становится отрицательной величиной, так как при циторризе действуют силы упругости клеточной оболочки, которые в увядших клетках растягивают протопласт, а не сдавливают его. Поэтому сосущая сила таких клеток равна сумме давлений осмотического и тургор ного, т. е. S = P + T, так как в этом случае S = P — (-Т).
Наблюдениями В. С. Шардакова над хлопчатником было установлено, что сила циторриза — сила растяжения протопласта клеточной оболочкой — может достигать 1010-1515 кПа
Сосущую силу клеток можно определить несколькими методами, например измерением длины узких полосок растительной ткани. Продольные вырезки тканей измеряют, затем помещают в растворы различной концентрации и через определенный промежуток времени снова измеряют. Если длина ткани уменьшилась, то внешний раствор был гипертоническим, а если увеличилась — гипотоническим. Нужно найти изотонический раствор, при котором размер ткани не изменяется. Концентрация этого раствора и будет соответствовать сосущей силе клеток растительной ткани. Достаточно точным является метод струек, предложенный В. С. Шардаковым, который используют в исследованиях по засухоустойчивости и орошению.
Абсолютная величина осмотического давления в клетках живых растений достаточно высокая и колеблется чаще всего от 506 до 1010 кПа у наземных растений и от 101 до 304 кПа у водных, В плодах фруктовых деревьев, ягодах винограда, корнеплодах сахарной свеклы осмотическое давление бывает в пределах 2026-4052 кПа, у растений, которые выращивают на засоленных почвах, оно может достигать 6078-10130 кПа. Величина сосущей силы в разных органах и тканях имеет важное значение для поступления и движения воды по растению.
Наблюдается колебание уровня сосущей силы. Например, величина этого показателя у листьев бука, взятых на разной высоте, неодинакова: чем выше расположены листья, тем она больше. Такое распределение величины сосущей силы по стволу дерева имеет большое значение для движения воды в нем.
Величины осмотического давления в тканях листа располагаются в таком убывающем порядке: палисадная паренхима, губчатая паренхима, верхний и нижний эпидермис. Клетки молодых листьев, как правило, имеют более высокую осмотическую концентрацию клеточного сока, чем клетки старых листьев. Вот почему иногда при недостатке воды, особенно в конце лета, нижние листья засыхают. Это происходит в результате оттягивания воды от них верхними листьями.
Величина осмотической концентрации клеточного сока имеет приспособительный характер. Например, у ячменя, прораставшего на охлажденной почве, осмотическое давление возрастало по сравнению с контрольными растениями, которые выращивали в нормальных условиях: в надземных органах — на 25, а в тканях корней — на 115%.
Физиологическое значение осмотических процессов велико. С ними связано поглощение растением воды и растворенных в ней минеральных веществ. Осмотическое давление в некоторой мере является регулятором движения воды по растению и распределения ее между отдельными органами. Оно имеет большое значение для обмена пластическими веществами между отдельными органами и тканями растений. В тканях дозревающих плодов наблюдается увеличение осмотического давления. Разностороннюю роль осмотической концентрации клеточного сока следует рассматривать как приспособительный механизм растений, выполняющий важную роль в обеспечении их водой и питательными веществами.
Источник