- Вода глюкоза мембрана процесс
- Вода глюкоза мембрана процесс
- Что такое фотосинтез и почему он так важен для нашей планеты
- Что такое фотосинтез
- Строение хлоропластов
- Пигменты хлоропластов
- Хлорофилл
- Каротиноиды
- Что происходит в процессе фотосинтеза
- Световая фаза фотосинтеза
- Темновая фаза фотосинтеза
- Значение фотосинтеза
- Заключение
Вода глюкоза мембрана процесс
• Для того чтобы преодолеть гематоэнцефалический барьер, глюкоза транспортируется в астроциты через эндотелиальные клетки небольших кровеносных сосудов
• Белки переносят глюкозу за счет унипорта, транспортируя ее в направлении градиента концентрации
• Белки, переносящие глюкозу, претерпевают конформационные изменения, которые приводят к реориентации их субстратных сайтов в мембране клетки
В клетках эукариот основным источником энергии является глюкоза, и многие клетки нуждаются в постоянном ее поступлении, поскольку у них глюкоза служит основным источником энергии для синтеза АТФ. Глюкоза представляет собой полярную молекулу, которая способна к гидратации, а клеточные мембраны не пропускают такие небольшие полярные метаболиты, как сахара. Поэтому для транспорта глюкозы в клетку требуется участие специфических мембранных белков. Транспорт глюкозы через плазматическую мембрану происходит с участием продуктов двух семейств генов.
Переносчики глюкозы (GLUTs) осуществляют независимый перенос (унипорт), при котором через мембрану происходит облегченный транспорт глюкозы. В противоположность GLUT-белкам, котранспортеры Na+/глюкоза расходуют энергию трансмембранного градиента Na+ на транспорт глюкозы. В настоящем разделе мы рассмотрим GLUT-белки.
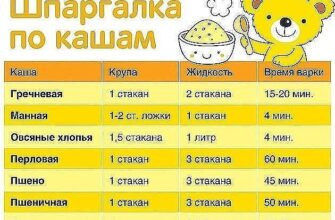
![]() Глюкоза селективно транспортируется через гематоэнцефалический барьер посредством изоформы 1 (GLUT-1) белка-переносчика.
Глюкоза селективно транспортируется через гематоэнцефалический барьер посредством изоформы 1 (GLUT-1) белка-переносчика.
Перенос глюкозы из крови в головной мозг и в другие отделы ЦНС представляет собой многоступенчатый процесс,
в котором участвуют различные типы клеток.
Семейство GLUT-белков является частью более обширного суперсемейства белков, облегчающих основные транспортные процессы (MFS) и широко представленных в клетках всех организмов. GLUT-белки представляют собой интегральные белки мембран клеток эукариот. Изоформы переносчиков GLUT различаются по своим кинетическим характеристикам, специфичностью по отношению к переносимым сахарам, тканевой локализацией и механизмами регуляции. Некоторые GLUT-белки, наряду с глюкозой, транспортируют и другие метаболиты, например галактозу, воду и анальгетики из группы гликопептидов.
GLUT-белки осуществляют унипорт, при котором вещества проходят через мембрану в направлении градиента концентрации. Таким образом, в зависимости от концентрации, GLUT-белки переносят метаболиты в клетку или в противоположном направлении. Поступление в клетку глюкозы, происходящее при участии GLUT-белков, часто определяет жизнеспособность клеток, которые характеризуются высоким уровнем потребления энергии.
Питательные метаболиты, такие как сахара, через кровеносные сосуды транспортируются в органы. Эндотелиальные клетки, выстилающие стенки небольших сосудов, контролируют процессы обмена питательных веществ. В этих эндотелиальных клетках, особенно расположенных в области гематоэнцефалического барьера, содержится много GLUT-белков. Надлежащее функционирование головного мозга сильно зависит от глюкозы, и его клетки особенно чувствительны к снижению ее содержания.
Транспорт глюкозы в нервные клетки происходит через капилляры мозга в несколько этапов и с участием изоформы GLUT-1. Эта изоформа экспрессируется в мембране клеток эндотелия, находящихся на границе между кровью и межклеточным пространством, а также в плазматической мембране астроцитов, функция которых важна в гематоэнцефалическом барьере. Расположенные в этих местах белки GLUT-1 транспортируют глюкозу из крови в эндотелиальные клетки, и оттуда в астроциты. В них глюкоза превращается в другие источники энергии, которые транспортируются в нейроны.
В различных тканях содержатся разные изоформы GLUT. Например, в клетках мышечной и жировой ткани транспорт глюкозы осуществляется с участием GLUT-4. В процессе приема пищи или после еды в клетки этих тканей под действием инсулина увеличивается поступление глюкозы. При этом происходит регулируемый транспорт изоформы GLUT-4 к клеточной поверхности. Эта изоформа также называется инсулин-зависимый переносчик. Белок GLUT-4 локализован во внутриклеточных везикулах, которые сливаются с плазматической мембраной. Этим обеспечивается доставка переносчика GLUT-4 к плазматической мембране и увеличивается емкость транспортного процесса.
![]() Предполагаемое строение GLUT-переносчика, состоящего из 12 трансмембранных сегментов с внутриклеточными С- и N-концевыми участками.
Предполагаемое строение GLUT-переносчика, состоящего из 12 трансмембранных сегментов с внутриклеточными С- и N-концевыми участками.
Внутриклеточные петли содержат сайты фосфорилирования и связывания субстратов.
Показано, что градиент глюкозы по обеим сторонам мембраны может возникать в любом направлении, в зависимости от типа клеток и их метаболизма.
Градиент определяет направление транспорта.
Связывание инсулина со своим поверхностным рецептором запускает каскад внутриклеточных процессов, приводящих к быстрому слиянию этих везикул с мембраной. Это, в свою очередь, приводит к быстрому увеличению транспорта глюкозы в клетку с участием GLUT-4. При диабете типа II не происходит транспорт глюкозы из крови в мышцы и жировую ткань, очевидно, из-за нарушения позиционирования GLUT-4 на плазматической мембране. Более того, GLUT-2 способен экспортировать глюкозу из клеток тех органов, в которых она образуется, например из клеток печени.
По строению переносчик GLUT напоминает другие члены суперсемейства MFS. Предполагается, что он состоит из 12 трансмембранных а-спиралей, имеющих внутриклеточные N- и С-концевые участки и петли. Последние содержат сайт связывания субстрата и сайты фосфорилирования.
Модель структуры GLUT-1 предложена на основании результатов по сайт-направленному мутагенезу и по измерению транспорта глюкозы мутантным белком. Для построения модели также использовались данные по кристаллической структуре бактериальной лактопермеазы, относящейся к подсемейству переносчиков олигосахари-дов/Н+, входящих в MFS.
Согласно этой модели, при соответствующей ориентации трансмембранных спиралей создается полость поры, через которую транспортируется глюкоза, а также обеспечивается образование водородных связей между GLUT-белком и глюкозой. Результаты кинетического анализа транспорта глюкозы в эритроцитах позволяют предполагать существование специального механизма, который обеспечивает белку GLUT возможность принимать одну из двух основных конформаций. Этот механизм аналогичен постулируемому для бактериальной лактопермеазы. Когда белок находится в одной конформации, сайт связывания глюкозы обращен в сторону внеклеточного пространства. В другой конформации он обращен в сторону цитозоля. При связывании глюкозы с любой стороны происходят конформационные изменения, которые приводят к реориентации сайтов связывания глюкозы к противоположной стороне мембраны и к ее высвобождению.
Таким образом, хотя переносчики GLUT являются унипортерами, а бактериальная лактопермеаза представляет собой симпортер, предполагается, что они функционируют по одному механизму.
Мутации в гене GLUT-1 вызывают значительные дефекты развития. У детей потребность головного мозга в глюкозе в 3-4 раза выше, чем у взрослых, и в мозг поступает до 80% всей глюкозы. Мутации в гене GLUT-1 человека служат причиной проявления редкого синдрома недостаточности GLUT-1, который характеризуется развитием припадков и замедлением развития. Считается, что это связано с нарушением транспорта глюкозы в головной мозг. Эмбрионы мышей, дефектных по гену GLUT-1, отстают в росте, и у них обнаруживаются различные пороки развития. Аналогичные дефекты возникают у мышиных эмбрионов, которые развиваются в организме диабетических самок. При этом избыток глюкозы в крови подавляет экспрессию GLUT-1 в органах эмбриона.
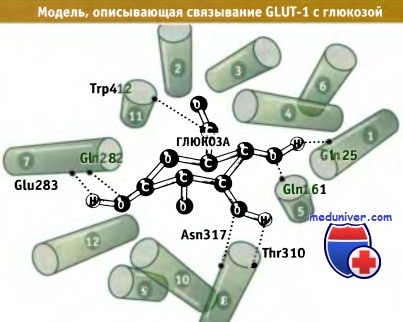 Предполагаемая ориентация трансмембранных а-спиралей переносчика глюкозы GLUT-1.
Предполагаемая ориентация трансмембранных а-спиралей переносчика глюкозы GLUT-1.
Показаны остатки, участвующие в связывании глюкозы (размер молекулы глюкозы представлен без учета масштаба).
Вид на спирали со стороны внутренней части плазматической мембраны.
Эта модель построена с использованием гомологичной структуры лактопермеазы E.coli в качестве матрицы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Вода глюкоза мембрана процесс
а) Главенствующая роль глюкозы в метаболизме углеводов. Конечными продуктами гидролиза углеводов в желудочно-кишечном тракте являются всего три вещества: глюкоза, фруктоза и галактоза. При этом на долю глюкозы приходится почти 80% общего количества этих моносахаридов. После всасывания в кишечнике большая часть фруктозы и практически вся галактоза преобразуются в печени в глюкозу. Вследствие этого в крови присутствуют только небольшие количества фруктозы и галактозы. В итоге процессов превращения глюкоза становится единственным представителем углеводов, транспортируемым во все клетки организма.
Соответствующие ферменты, необходимые клеткам печени для обеспечения процессов взаимного превращения моносахаридов — глюкозы, фруктозы и галактозы — показаны на рисунке ниже.
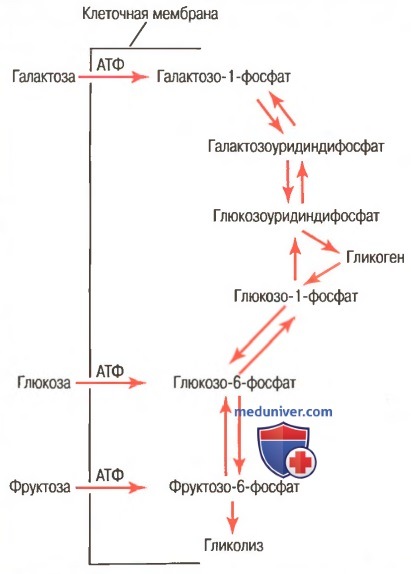 Взаимные превращения трех основных моносахаридов (глюкозы, фруктозы и галактозы) в клетках печени
Взаимные превращения трех основных моносахаридов (глюкозы, фруктозы и галактозы) в клетках печени
В результате этих реакций, когда печень высвобождает моносахариды обратно в кровь, окончательным продуктом, попадающим в кровь, становится глюкоза. Причина этого явления заключается в том, что клетки печени содержат большое количество глюкозофосфатазы, поэтому глюкозо-6-фосфат может расщепляться на глюкозу и фосфат. Затем глюкоза транспортируется через мембраны клеток обратно в кровь.
Хотелось бы еще раз подчеркнуть, что обычно более 95% всех моносахаридов, циркулирующих в крови, представлены конечным продуктом превращения — глюкозой.
б) Транспорт глюкозы через мембрану клетки. Прежде чем глюкоза будет использована клетками тканей, она должна транспортироваться через мембраны клеток в цитоплазму. Однако глюкоза не может свободно диффундировать через поры в клеточных мембранах, т.к. максимальная молекулярная масса частиц должна быть в среднем равна 100, в то время как молекулярная масса глюкозы составляет 180. Тем не менее глюкоза может относительно легко проникать внутрь клеток благодаря механизму облегченной диффузии. Основы этого механизма обсуждались в главе 4, напомним его основные моменты.
Насквозь прободая липидную мембрану клеток, белки-переносчики, количество которых в мембране достаточно велико, могут взаимодействовать с глюкозой. В такой связанной форме глюкоза может транспортироваться белком-переносчиком с одной стороны мембраны на другую и там отделяться; если с одной стороны мембраны концентрация глюкозы выше, чем с другой, то глюкоза будет транспортироваться туда, где ее концентрация ниже, а не в противоположном направлении. Транспорт глюкозы через клеточные мембраны в большинстве тканей резко отличается от транспорта, который наблюдается в желудочно-кишечном тракте или в эпителиоцитах канальцевого аппарата почек.
В обоих упомянутых случаях транспорт глюкозы опосредован сопряженным с механизмом активного транспортом натрия. Активный транспорт натрия обеспечивает энергией процесс всасывания глюкозы против градиента концентрации. Такой сопряженный с натрием активный механизм транспорта глюкозы встречается только в специализированных эпителиоцитах, приспособленных для активного процесса абсорбции глюкозы. В других клеточных мембранах глюкоза транспортируется только из областей с высокой концентрацией в область низких концентраций с помощью механизма облегченной диффузии, возможность которого создается особыми свойствами расположенного в мембране белка-переносчика глюкозы.
Подробно механизм облегченной диффузии применительно к транспорту через клеточные мембраны изложен в отдельной статье на сайте (просим вас пользоваться формой поиска выше).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Что такое фотосинтез и почему он так важен для нашей планеты
Фотосинтез — один из самых важных биологических процессов на Земле. Благодаря фотосинтезу живые организмы получают кислород, необходимый для дыхания, а сами растения создают полезные органические вещества для своей жизнедеятельности. В этой статье мы поговорим о том, что обозначает фотосинтез, как он происходит и что образуется в процессе фотосинтеза.

Что такое фотосинтез
Фотосинтез — процесс, при котором в клетках, содержащих хлорофилл, под действием энергии света образуются органические вещества из неорганических. При фотосинтезе растение поглощает углекислый газ и воду, синтезирует органические вещества и выделяет кислород, как побочный продукт фотосинтеза.
Процессы фотосинтеза идут в тканях, содержащих хлоропласты, — преимущественно, в листе, на который приходится большая часть процессов фотосинтеза. Такая ткань называется хлоренхима, или мезофилл.
Строение хлоропластов
Чтобы понять, что происходит в растении при фотосинтезе, изучим подробнее хлоропласты. Хлоропласты — это особые пластиды растительных клеток, в которых происходит фотосинтез. Основные элементы структурной организации хлоропластов высших растений представлены на рис.1.

Хлоропласт — это двумембранный органоид. Внешняя мембрана проницаема для большинства органических и неорганических соединений. Она содержит специальные транспортные белки, благодаря которым нужные для работы хлоропласта пептиды и другие вещества попадают в него из цитоплазмы. Внутренняя мембрана обладает избирательной проницаемостью и способна контролировать, какие именно вещества попадут во внутреннее пространство хлоропласта.

Для хлоропластов характерна сложная система внутренних мембран, позволяющая пространственно организовать фотосинтетический аппарат, упорядочить и разделить реакции фотосинтеза, несовместимые между собой, и их продукты. Мембраны образуют тилакоиды, которые, в свою очередь, собираются в «стопки» — граны. Пространство внутри тилакоидов называется внутритилакоидным пространством, или люменом.
Внутреннее пространство хлоропласта между гранами заполняет строма — гидрофильный слабоструктурированный матрикс. В строме содержатся необходимые для реакций синтеза сахаров ферменты, а также рибосомы, кольцевая молекула ДНК, крахмальные зёрна.
Пигменты хлоропластов
Что происходит во время фотосинтеза? На молекулярном уровне фотосинтез обеспечивают особые вещества — пигменты, благодаря которым энергия солнечного света становится доступной для биологических систем. У фотосинтезирующих организмов можно выделить три основные группы пигментов:
- Хлорофиллы:
- хлорофилл а — у большинства фотосинтезирующих организмов,
- хлорофилл b — у высших растений и зелёных водорослей,
- хлорофилл c — у бурых водорослей,
- хлорофилл d — у некоторых красных водорослей.
- Каротиноиды:
- каротины — у всех фотосинтезирующих организмов, кроме прокариот;
- ксантофиллы — у всех фотосинтезирующих организмов, кроме прокариот
- Фикобилины — красные и синие пигменты красных водорослей.
В хлоропластах пигменты ассоциированы с белками с помощью ионных, водородных и других типов связей. Не стоит забывать, что у растений есть множество других пигментов, находящихся не в хлоропластах и не принимающих участие в фотосинтезе — например, антоцианы.
Хлорофилл
Хлорофиллы выполняют функции поглощения, преобразования и транспорта энергии света. Лучше всего хлорофиллы поглощают свет в синей (430—460 нм) и красной (650—700 нм) областях спектра. Зелёную область спектра хлорофиллы эффективно отражают, что придаёт растению зелёный цвет.
Интересно, что строение молекулы хлорофилла схоже со строением гемоглобина, но центром молекулы хлорофилла является ион магния, а не железа.
Основными хлорофиллами высших растений являются хлорофилл a и хлорофилл b, они входят в состав реакционных центров фотосистем и светособирающих комплексов мембран тилакоидов хлоропластов. Светособирающие комплексы улавливают кванты света и передают энергию к фотосистемам I и II. Фотосистемы — это пигмент-белковые комплексы, играющие ключевую роль в световой фазе фотосинтеза.
Каротиноиды
Каротиноиды — это жёлтые, оранжевые или красные пигменты. В зелёных листьях каротиноиды обычно незаметны из-за наличия в листьях хлорофилла. При разрушении хлорофилла осенью именно каротиноиды придают листьям характерную жёлто-оранжевую окраску.
- Антенная — входят в состав светособирающих комплексов, улавливают энергию света и передают её на хлорофиллы. Каротиноиды играют роль дополнительных светособирающих пигментов в той части солнечного спектра (450—570 нм), где хлорофиллы малоэффективны. Особенно это важно для водных экосистем, в которых волны оптимальной для хлорофиллов длины быстро исчезают с глубиной.
- Защитная функция (антиоксидантная) — обезвреживание агрессивных кислородных соединений (активных форм кислорода) и избытка хлорофилла в возбуждённом состоянии при слишком ярком освещении.
Каротиноиды химически представляют собой 40-углеродную цепь с двумя углеродными кольцами по краям цепи. В строении ксантофиллов, в отличие от каротинов, присутствуют спиртовые, эфирные или альдегидные группы.
Учите биологию вместе с домашней онлайн-школой «Фоксфорда»! По промокоду BIO72021 вы получите бесплатный доступ к курсу биологии 7 класса, в котором изучается тема фотосинтеза.
Что происходит в процессе фотосинтеза
Как уже было сказано ранее, в ходе фотосинтеза в хлоропластах под действием солнечного света образуются органические вещества.
Процесс фотосинтеза можно разделить на две фазы:
В ходе световой фазы фотосинтеза образуется энергия в виде АТФ и универсальный донор атома водорода — восстановитель НАДФН (НАДФ·Н2). Эти вещества необходимы для протекания темновой фазы. Также образуется побочный продукт — кислород. Световая фаза может проходить только на мембранах тилакоидов и на свету.
Благодаря сложному биохимическому процессу — циклу Кальвина — в темновую фазу фотосинтеза образуются органические вещества (сахара). Темновая фаза проходит в строме хлоропластов и на свету, и в темноте. Темновые ферментативные процессы протекают медленнее, чем световые, поэтому при очень ярком освещении скорость протекания фотосинтеза будет полностью определяться скоростью темновой фазы. Схемы процессов фотосинтеза представлены на рис.2. Подробное описание процессов смотри далее.

Световая фаза фотосинтеза
Чтобы лучше понять, что происходит во время фотосинтеза, разберём фазы фотосинтеза. Световая фаза фотосинтеза включает в себя фотохимические и фотофизические процессы, и может быть поделена на три этапа:
- Фаза поглощения — энергия света улавливается при помощи светособирающих комплексов, переходит в энергию электронного возбуждения пигментов, передаётся в реакционный центр фотосистем I и II.
- Фаза реакционных центров — энергия электронного возбуждения пигментов светособирающих комплексов используется для активации реакционных центров фотосистем. В реакционном центре электрон от возбуждённого хлорофилла передаётся другим компонентам электрон-транспортной цепи, пигмент после отдачи электрона переходит в окисленное состояние и становится способным, в свою очередь, отнимать электроны у других веществ. Именно в этом процессе происходит преобразование физической формы энергии в химическую.
- Фаза электрон-транспортной цепи — электроны переносятся по цепи переносчиков, образуются АТФ, НАДФН, O2. Необходимо, чтобы каждый переносчик электрон-транспортной цепи поочерёдно восстанавливался и окислялся, обеспечивая таким образом перенос энергии электронов. Любой этап переноса электрона сопровождается высвобождением или поглощением энергии. Часть энергии теряется. На некоторых участках электрон-транспортной цепи перенос электрона сопряжён с переносом протона.
Для того чтобы понять, что происходит во время фазы фотосинтеза, рассмотрим эти процессы подробнее. Кванты света улавливаются светособирающими комплексами фотосистемы I — молекула хлорофилла в составе светособирающего комплекса переходит в возбуждённое состояние, и энергия передаётся в реакционный центр фотосистемы I. Происходит возбуждение молекул хлорофилла фотосистемы I, отщепляется электрон. Пройдя по цепочке внутренних компонентов фотосистемы I и внешних переносчиков, электрон в конце концов попадает к НАДФ+ — образуется восстановитель НАДФН. Получается, что хлорофилл фотосистемы I отдал электрон и приобрёл положительный заряд, и для дальнейшего функционирования необходимо восстановить нейтральность молекулы, получить электрон, чтобы закрыть «дырку». Этот электрон приходит от фотосистемы II.
На светособирающие комплексы фотосистемы II попадают кванты света — происходит возбуждение молекулы хлорофилла фотосистемы II, молекула хлорофилла отдаёт электрон и переходит в окисленное состояние. Нехватку электрона хлорофилл восполняет благодаря фотолизу воды, при этом образуется протоны H+, а также важный побочный продукт фотосинтеза — кислород. По цепи переносчиков электрон от хлорофилла фотосистемы II попадает к хлорофиллу реакционного центра фотосистемы I и восстанавливает его. Теперь этот хлорофилл может снова поглощать энергию кванта света и отдавать электрон в электрон-транспортную цепь.
Протоны, попадающие во внутритилакоидное пространство, используются для синтеза АТФ. С помощью фермента АТФ-синтазы за счёт градиента протонов образуется АТФ из АДФ и фосфата. Под градиентом понимают неравномерное распределение: во внутритилакоидном пространстве H+ больше, в строме — меньше. Поэтому частицы стремятся проникнуть в строму, переходят в неё через АТФ-синтазу, а в процессе пути сквозь белковый комплекс отдают ему часть энергии, которая и используется для синтеза АТФ.
Темновая фаза фотосинтеза
Что образуется при фотосинтезе в темновую фазу? В строме хлоропластов с помощью энергии АТФ и восстановителя НАДФН, полученных в световую фазу, образуются простые сахара, из которых в ходе других процессов образуется крахмал. Ферментативные процессы не нуждаются в наличии света. Важнейший процесс, происходящий в темновую фазу фотосинтеза, — фиксация углекислого газа воздуха. Синтез и превращения сахаров в хлоропластах имеют циклический характер и носят название цикл Кальвина.
В нём можно выделить три этапа:
- Фаза карбоксилирования (введение CO2 в цикл).
- Фаза восстановления (используются АТФ и НАДФН, полученные в световую фазу).
- Фаза регенерации (превращения сахаров).
В строме хлоропластов находится производное простого пятиуглеродного сахара рибозы. С помощью особого фермента (Рубиско) к производному рибозы присоединяется CO2 (реакция карбоксилирования) — образуется неустойчивое шестиуглеродное соединение, которое быстро распадается на две трехуглеродные молекулы. Дальше, с затратой АТФ и НАДФН, полученных в ходе световых процессов, трехуглеродное соединение модифицируется — образуется восстановленное соединение с атомом фосфора и альдегидной группой в составе. Теперь перед клеткой стоит проблема: необходимо получить шестиуглеродное соединение — глюкозу для синтеза крахмала, а также пятиуглеродное — производное рибозы для того, чтобы эти процессы могли начаться заново. Для решения этих проблем в фазу регенерации из полученных ранее трехуглеродных соединений под действием ферментов образуются четырёх-, пяти-, шести- и семиуглеродные сахара. Из шестиуглеродной молекулы образуется глюкоза, из которой синтезируется крахмал. Из пятиуглеродной молекулы образуется производное рибозы и цикл замыкается. Остальные сахара также используются клеткой в других биохимических процессах.
Отдельно стоит сказать про крайне важный фермент первой фазы цикла Кальвина — рибулозо-1,5-дифосфаткарбоксилазу (Рубиско). Это сложный фермент, состоящий из 16 субъединиц, с молекулярной массой в 8 раз больше, чем у гемоглобина. Является одним из важнейших ферментов в природе, поскольку играет центральную роль в основном механизме поступления неорганического углерода (из CO2) в биологический круговорот. Содержание Рубиско в листьях растений очень велико, он считается самым распространённым ферментом на Земле.

Значение фотосинтеза
В процессе фотосинтеза энергия света заключается в энергию химических связей органических веществ. Поэтому фотосинтез служит первичным источником почти всей энергии, используемой живыми организмами в процессе жизнедеятельности. Практически все живые организмы, за исключением хемосинтетиков, так или иначе пользуются теми продуктами, что выделяются при фотосинтезе.
За счёт фотосинтеза сформировалась и поддерживается пригодная для дыхания атмосфера с высоким содержанием кислорода.
Фиксация углекислого газа в ходе фотосинтеза служит главным местом входа неорганического углерода в биогеохимический цикл. Также ассимиляция CO2 препятствует перегреву Земли, предотвращая парниковый эффект.
Заключение
Каждый год на нашей планете благодаря фотосинтезу производится около 200 миллиардов тонн кислорода, из которого образуется озоновый слой, защищающий от ультрафиолетовой радиации. Фотосинтез помогает поддерживать состав атмосферы и препятствует увеличению количества углекислого газа. Без растений и кислорода, который они выделяют в процессе фотосинтеза, жизнь на нашей планете была бы просто невозможна.
Источник