Глава 2. Гидросфера
2.1. Вода как среда обитания
В процессе исторического развития живые организмы освоили четыре среды обитания. Первая – вода. В воде жизнь зародилась и развивалась многие миллионы лет. Вторая – наземно-воздушная – на суше и в атмосфере возникли и бурно адаптировались к новым условиям растения и животные. Постепенно преобразуя верхний слой суши – литосферы, они создали третью среду обитания – почву, а сами стали четвертой средой обитания.
Вода покрывает 71 % площади земного шара и составляет 1/800 часть объема суши. Основная масса воды сосредоточена в морях и океанах – 94–98 %, в полярных льдах содержится около 1,2 % воды и совсем малая доля – менее 0,5 %, в пресных водах рек, озер и болот. Соотношения эти постоянны, хотя в природе, не переставая, идет круговорот воды.
В водной среде обитает около 150 000 видов животных и
10 000 растений, что составляет соответственно всего 7 и 8 % от общего числа видов Земли. В Мировом океане, как в горах, выражена вертикальная зональность. Особенно сильно различаются по экологии пелагиаль – вся толща воды, и бенталь – дно.
Особенно чётко зональность проявляется в озёрах умеренных широт (рис. 2.1). В водной массе как среде обитания организмов по вертикали может быть выделено 3 слоя: эпилимнион, металимнион и гиполимнион. Воды поверхностного слоя – эпилимниона летом прогреваются и перемешиваются под воздействием ветра и конвекционных токов. Осенью поверхностные воды, охлаждаясь и становясь более плотными, начинают погружаться, и температурная разность слоев выравнивается. При дальнейшем охлаждении воды эпилимниона становятся холоднее вод гиполимниона. Весной происходит обратный процесс, заканчивающийся периодом летнего застоя. Дно озёр (бенталь) подразделяется на 2 зоны: более глубоководную – профундаль, примерно соответствующую части ложа, заполненной водами гиполимниона, и прибрежную зону – литораль, обычно простирающуюся вглубь до границы произрастания макрофитов. По поперечному профилю реки различают прибрежную зону – рипаль и открытую – медиаль. В открытой зоне скорости течения выше, население количественно беднее, чем в прибрежной.
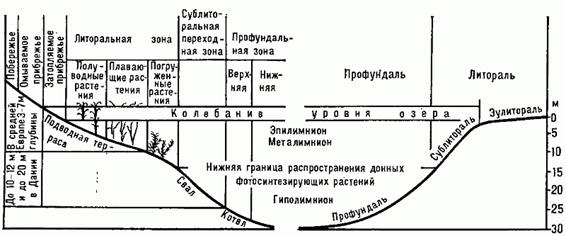
Рис. 2.1. Вертикальная экологическая зональность озера
(по М.С. Гилярову)
Экологические группы гидробионтов. Наибольшим разнообразием жизни отличаются теплые моря и океаны (40000 видов животных) в области экватора и тропиках, к северу и югу происходит обеднение флоры и фауны морей в сотни раз. Что касается распределения организмов непосредственно в море, то основная масса их сосредоточена в поверхностных слоях (эпипелагиаль) и в сублиторальной зоне. В зависимости от способа передвижения и пребывания в определенных слоях, морские обитатели подразделяются на три экологические группы: нектон, планктон и бентос.
Нектон (nektos – плавающий) – активно передвигающиеся крупные животные, способные преодолевать большие расстояния и сильные течения: рыбы, кальмары, ластоногие, киты. В пресных водоемах к нектону относятся и земноводные и множество насекомых.
Планктон (planktos – блуждающий, парящий) – совокупность растений (фитопланктон: диатомовые, зеленые и сине-зеленые (только пресные водоемы) водоросли, растительные жгутиконосцы, перидинеи и др.) и мелких животных организмов (зоопланктон: мелкие ракообразные, из более крупных – крылоногие моллюски, медузы, гребневики, некоторые черви), обитающих на разной глубине, но не способных к активным передвижениям и к противостоянию течениям. В состав планктона входят и личинки животных, образуя особую группу – нейстон. Это пассивно плавающее «временное» население самого верхнего слоя воды, представленное разными животными (десятиногие, усоногие и веслоногие ракообразные, иглокожие, полихеты, рыбы, моллюски и др.) в личиночной стадии. Личинки, взрослея, переходят в нижние слои пелагели. Выше нейстона располагается плейстон – это организмы, у которых верхняя часть тела растет над водой, а нижняя – в воде (ряска, кубышки, кувшинки и др.). Планктон играет важную роль в трофических связях биосферы, т.к. является пищей для многих водных обитателей, в том числе основным кормом для усатых китов.
Бентос (benthos – глубина) – гидробионты дна. Представлен в основном прикрепленными или медленно передвигающимися животными (зообентос: фораминефоры, рыбы, губки, кишечнополостные, черви, плеченогие моллюски, асцидии, и др.), более многочисленными на мелководье. На мелководье в бентос входят и растения (фитобентос: диатомовые, зеленые, бурые, красные водоросли, бактерии). На глубине, где нет света, фитобентос отсутствует. У побережий встречаются цветковые растения зостера, рупия. Наиболее богаты фитобентосом каменистые участки дна.
В озерах зообентос менее обилен и разнообразен, чем в море. Его образуют простейшие (инфузории, дафнии), пиявки, моллюски, личинки насекомых и др. Фитобентос озер образован свободно плавающими диатомеями, зелеными и сине-зелеными водорослями; бурые и красные водоросли отсутствуют.
Укореняющиеся прибрежные растения в озерах образуют четко выраженные пояса, видовой состав и облик которых согласуются с условиями среды в пограничной зоне «суша-вода». В воде у самого берега растут гидрофиты – полупогруженные в воду растения (стрелолист, белокрыльник, камыши, рогоз, осоки, трищетинник, тростник). Они сменяются гидатофитами – растениями, погруженными в воду, но с плавающими листьями (лотос, ряски, кубышки, чилим, такла) и – далее – полностью погруженными (рдесты, элодея, хара). К гидатофитам относятся и плавающие на поверхности растения (ряска).
Высокая плотность водной среды определяет особый состав и характер изменения жизнеобеспечивающих факторов. Одни из них те же, что и на суше – тепло, свет, другие специфические: давление воды (с глубиной увеличивается на 1 атм. на каждые 10 м), содержание кислорода, состав солей, кислотность. Благодаря высокой плотности среды, значения тепла и света с градиентом высоты изменяются гораздо быстрее, чем на суше.
Тепловой режим. Для водной среды характерен меньший приход тепла, т.к. значительная часть его отражается, и не менее значительная часть расходуется на испарение. Согласуясь с динамикой наземных температур, температура воды обладает меньшими колебаниями суточных и сезонных температур. Более того, водоемы существенно выравнивают ход температур в атмосфере прибрежных районов. При отсутствии ледового панциря моря в холодное время года оказывают отепляющее действие на прилегающие территории суши, летом – охлаждающее и увлажняющее.
Диапазон значений температуры воды в Мировом океане составляет 38° (от –2 до +36°С), в пресных водоемах – 26° (от –0,9 до +25°С). С глубиной температура воды резко падает. До 50 м наблюдаются суточные колебания температуры, до 400 – сезонные, глубже она становится постоянной, опускаясь до +1–3°С (в Заполярье близка к 0°С). Поскольку температурный режим в водоемах сравнительно стабилен, их обитателям свойственна стенотермность. Незначительные колебания температуры в ту или иную сторону сопровождается существенными изменениями в водных экосистемах.
Примеры: «биологический взрыв» в дельте Волги из-за понижения уровня Каспийского моря – разрастание зарослей лотоса (Nelumba kaspium), в южном Приморье – зарастание белокрыльником стариц рек (Комаровка, Илистая и др.) по берегам которых вырублена и сожжена древесная растительность. Для Беларуси можно привести случаи разрастания в водоемах таких несвойственных для нашей природной зоны гидробионтов как водяная сосенка и водный гиацинт в некоторых водоемах Полесья.
В связи с разной степенью прогревания верхних и нижних слоев в течение года, приливами и отливами, течениями, штормами происходит постоянное перемешивание водных слоев. Роль перемешивания воды для водных обитателей (гидробионтов) исключительно велика, т.к. при этом выравнивается распределение кислорода и питательных веществ внутри водоемов, обеспечивая обменные процессы между организмами и средой.
В стоячих водоемах (озерах) умеренных широт весной и осенью имеет место вертикальное перемешивание, и в эти сезоны температура во всем водоеме становится однородной, т.е. наступает гомотермия. Летом и зимой в результате резкого усиления прогревания или охлаждения верхних слоев перемешивание воды прекращается. Это явление называется температурной дихотомией, а период временного застоя – стагнацией (летней или зимней). Летом более легкие теплые слои остаются на поверхности, располагаясь над тяжелыми холодными. Зимой, наоборот, в придонном слое более теплая вода, так как непосредственно подо льдом температура поверхностных вод меньше +4°С и они в силу физико-химических свойств воды становятся более легкими, чем вода с температурой выше +4°С.
В периоды стагнаций четко выделяются три слоя: верхний (эпилимнион) с наиболее резкими сезонными колебаниями температуры воды, средний (металимнион или термоклин), в котором происходит резкий скачок температур, и придонный (гиполимнион), в котором температура в течение года изменяется слабо. В периоды стагнаций в толще воды образуется дефицит кислорода – летом в придонной части, а зимой и в верхней, вследствие чего в зимний период нередко происходят заморы рыбы. В стоячих водоемах (озерах) умеренных широт весной и осенью имеет место вертикальное перемешивание, и в эти сезоны температура во всем водоеме становится однородной, т.е. наступает гомотермия. Летом и зимой в результате резкого усиления прогревания или охлаждения верхних слоев перемешивание воды прекращается. Это явление называется температурной дихотомией, а период временного застоя – стагнацией (летней или зимней). Летом более легкие теплые слои остаются на поверхности, располагаясь над тяжелыми холодными. Зимой, наоборот, в придонном слое более теплая вода, так как непосредственно подо льдом температура поверхностных вод меньше +4°С и они в силу физико-химических свойств воды становятся более легкими, чем вода с температурой выше +4°С.
В периоды стагнаций четко выделяются три слоя: верхний (эпилимнион) с наиболее резкими сезонными колебаниями температуры воды, средний (металимнион или термоклин), в котором происходит резкий скачок температур, и придонный (гиполимнион), в котором температура в течение года изменяется слабо. В периоды стагнаций в толще воды образуется дефицит кислорода – летом в придонной части, а зимой и в верхней, вследствие чего в зимний период нередко происходят заморы рыбы.
Световой режим. Интенсивность света в воде сильно ослаблена из-за его отражения поверхностью и поглощения самой водой. Это сильно сказывается на развитии фотосинтезирующих растений. Чем меньше прозрачность воды, тем сильнее поглощается свет. Прозрачность воды лимитируется минеральными взвесями, планктоном. Уменьшается она при бурном развитии мелких организмов летом, а в умеренных и северных широтах – еще и зимой, после установления ледового покрова и укрытия его сверху снегом.
В небольших озерах на глубину 2 м проникает всего лишь десятые доли процента света. С глубиной становится все темнее, и цвет воды становится вначале зеленым, затем голубым, синим и в конце – сине-фиолетовым, переходя в полный мрак.
Соответственно меняют цвет и гидробионты, адаптирующиеся не только к составу света, но и к его недостатку – хроматическая адаптация. В светлых зонах, на мелководьях, преобладают зеленые водоросли (Chlorophyta), хлорофилл которых поглощают красные лучи, c глубиной они сменяются бурыми (Phaephyta) и далее красными (Rhodophyta). На больших глубинах фитобентос отсутствует.
К недостатку света растения приспособились развитием хроматофоров крупных размеров, обеспечивающих низкую точку компенсации фотосинтеза, а также увеличением площади ассимилирующих органов (индекса листовой поверхности). Для глубоководных водорослей типичны сильно рассеченные листья, пластинки листьев тонкие, просвечивающиеся. Для полупогруженных и плавающих растений характерна гетерофиллия – листья над водой такие же, как у наземных растений, имеют цельную пластинку, развит устьичный аппарат, а в воде листья очень тонкие, состоят из узких нитевидных долей.
Животные, как и растения, закономерно меняют свою окраску с глубиной. В верхних слоях они ярко окрашены в разные цвета, в сумеречной зоне (морской окунь, кораллы, ракообразные) окрашены в цвета с красным оттенком – удобнее скрываться от врагов. Глубоководные виды лишены пигментов.
Источник
Вода как среда обитания




![]()
![]()
Как среда обитания организмов вода имеет ряд специфических свойств (большая плотность, значительные перепады давления, относительно малое содержание кислорода, высокая теплоемкость, сильное поглощение солнечных лучей и др.). Отдельные водоемы различаются по солевому режиму, по наличию течений и по др. параметрам.
Обитатели водной среды в экологии называются гидробионтами. По условиям обитания гидробионтов в любом водоеме можно выделить различные зоны. Рассмотрим экологические зоны мирового океана. В океане вместе с входящими в него морями можно выделить две основные экологические зоны: толщу воды – пелагиаль и дно – бенталь. Бенталь в зависимости от глубины подразделяется на зоны: сублитораль– область плавного увеличения глубины до 200 м; батиаль – крутой склон дна; абиссаль – океаническое ложе со средней глубиной 3–6 км; ультраабиссаль – впадины океанического ложа; литораль– кромка берега, заливаемая приливами; супралитораль – часть берега, увлажняемого брызгами прибоя.
Совокупность организмов, живущих на дне океана называется бентос, а живущих в толще воды – пелагос.
Пелагиаль также делится на зоны соответствующие зонам бентали: эпипелагиаль, батипелагиаль, абиссопелагиаль.Нижняя граница эпипелагиали (
200м) определяется количеством проникающего солнечного света, достаточным для фотосинтеза. Глубже этих зон зеленые растения не могут существовать. В сумеречных батиальных и полных мрака абиссальных глубинах обитают лишь микроорганизмы и животные.
Основные свойства водной среды.
Плотность воды – фактор, определяющий условия передвижения водных организмов и давление на разных глубинах. Максимальная плотность чистой (дистиллированной) воды достигается при температуре 4 о С и равна 1 г/см 3 . Природные воды имеют плотность до 1,35 г/см 3 . Давление растёт с глубиной в среднем на 1·10 5 Па (1 атм) на каждые 10м.
Гидробионты более эврибатны, нежели сухопутные организмы. Так, некоторые виды червей могут обитать и в прибрежной зоне и в ультраабиссали. Однако некоторые гидробионты стенобатны, т.е. обитают на строго определённых глубинах. Плотность воды обеспечивает возможность опираться на неё, что является необходимым условием парения в воде. Многие организмы приспособились именно к этому образу жизни и объединяются в особую экологическую группу гидробионтов – планктон. Планктон – это одноклеточные водоросли, простейшие, медузы, некоторые моллюски, мелкие рачки, личинки донных животных, икра и мальки рыб и др. Различают фитопланктон – водоросли и зоопланктон – остальное. Планктон не преодолевает течения и переносится ими на большие расстояния, однако некоторые его виды могут активно перемещаться по вертикали за счет регулирования плавучести тела.
Животных, способных к быстрому плаванию и преодолению сильных течений, объединяют в экологическую группу нектон – это рыбы, кальмары, дельфины. Их быстрое передвижение в воде возможно за счёт сильно развитой мускулатуры, сопряжённой с обтекаемой формой тела. Животные используют следующие основные способы движения в воде: реактивный; за счёт изгибания тела; с помощью конечностей.
Кислородный режим водоёмов.
В насыщенной кислородом воде содержание его составляет не менее 10 мл на 1 л, что в 21 раз меньше, чем в атмосфере. Кислород поступает в воду в основном за счёт фотосинтеза в водорослях и диффузии из воздуха. Поэтому верхние слои водоёмов богаче кислородом, чем нижние. С повышением температуры и солёности воды концентрация кислорода уменьшается. В слоях, сильно заселённых гидробионтами, может ощущаться дефицит кислорода из-за повышенного его потребления. Около дна водоёмов условия могут быть анаэробными.
Многие гидробионты эвриоксибионтны (сазан, линь, карась и др.); ряд видов стеноксибионтны и существуют лишь при высоком содержании кислорода (радужная форель, кумжа, гольян и др.). Многие виды способны впадать в состояние аноксибиоза при недостатке кислорода.
Дыхание гидробионтов осуществляется различными путями: поверхностью тела; жабрами; лёгкими; трахеями. При этом в ходе эволюции выработались различные адаптации, интенсифицирующие дыхание (утончение покровов тела и увеличение относительной его поверхности, перемешивание прилегающих к телу слоёв воды, комбинирование водного и воздушного дыхания и др.)

Недостаток кислорода приводит к заморам, сопровождающимся гибелью множества гидробионтов. Зимние заморы обусловлены образованием ледяного покрова, а летние повышением температуры воды и, как следствие, уменьшением растворимости кислорода в воде. Заморы, кроме недостатка кислорода, могут быть вызваны повышением концентрации в воде токсичных газов – метана (СН4), сероводорода (Н2S), диоксида серы (SО2), и др., образующихся в результате разложения органических веществ на дне водоемов.
Солевой режим водоёмов.
Особенностью гидробионтов является поддержание определенного количества воды в теле при её избытке в окружающей среде, т.к. изменение количества воды в клетках приводит к изменению в них осмотического давления и нарушению важнейших жизненных функций.
Водные обитатели в основном пойкилоосмотичны: осмотическое давление в их теле зависит от солёности окружающей воды, основной способ поддержания своего солевого баланса у них – выбор мест обитания с подходящей их потребностям солёностью: пресноводные не живут в морях, морские – не переносят опреснения. При изменении солёности воды животные мигрируют в поисках благоприятной среды. Так, например, при опреснении поверхностных слоёв моря после сильных дождей радиолярии, морские рачки и др. опускаются на глубину до 100 м.
Водные позвоночные, высшие раки, насекомые и их личинки являются гомойоосмотическими, сохраняя постоянное осмотическое давление в теле независимо от концентрации солей в воде.
Если соки тела гипертоничны по отношению к окружающей воде, то организмам угрожает обводнение, иначе – обезвоживание.
Защита организмов от этих неблагоприятных явлений обеспечивается различными путями: изменение концентрации солей в теле; наличие непроницаемых для воды покровов; переход в состояние солевого анабиоза.
Эвригалинных видов, способных в активном состоянии обитать как в пресной, так и в соленой воде немного, причем это в основном виды, населяющие дельты рек, лиманы и др. солоноватоводные водоемы.
Температурный режим водоемов.
Физические свойства воды обусловливают более устойчивый температурный режим по сравнению с наземно-воздушной средой по следующим причинам:
– высокая удельная теплоёмкость воды (4200 Дж/(кг∙К)), из-за которой получение или отдача значительного количества тепла не вызывает слишком резкого и большого изменения температуры водоёмов;
– высокая теплота парообразования воды (2,3·10 6 Дж/кг) при испарении её с поверхности водоёмов препятствует перегреванию последних;
– образование льда, которое происходит с выделением тепла (3,3∙10 5 Дж/кг), что замедляет охлаждение верхних слоёв.
Годовая амплитуда колебаний температуры в верхних слоях океана не > 10…15 о С, а в континентальных водоёмах
30…35 о С. В глубинных слоях водоёмов колебания температуры незначительны
Из-за более устойчивого температурного режима водоемов гидробионты более стенотермны, нежели наземные организмы.
Световой режим водоёмов:
В воде света гораздо меньше, чем в воздухе, т.к. происходит отражение его от поверхности воды и поглощение в её объеме. Изменение количества отражённого света в зависимости от высоты стояния солнца над горизонтом уменьшает продолжительность дня в водоёмах. Так, на глубинах
30 м продолжительность дня летом
5 час, а на глубине 40 м –
15 мин. Быстрое убывание количества света с глубиной обусловлено его поглощением массой воды. Разные длины волн оптического диапазона электромагнитного излучения солнца поглощаются неодинаково: сначала длинноволновые (красный цвет поглощается поверхностным слоем), затем все более коротковолновые лучи (сине-фиолетовые лучи достигают глубины
200 м при хорошей прозрачности воды).
Водоросли в Мировом океане обитают на глубине до 20…40 м, причем глубже других проникают красные водоросли (иногда до 100…200 м).
С глубиной меняется окраска животных: наиболее разнообразные цвета их в литоральной и сублиторальной зонах; в сумеречных глубинах превалирует красная окраска, являющаяся дополнительной к сине-фиолетовому цвету. Дополнительные по цвету лучи наиболее полно поглощаются телом животных, что позволяет им скрываться от врагов, т.к. их красный цвет в сине-фиолетовых лучах зрительно воспринимается как черный (морской окунь, красный коралл и др.). Глубинные организмы не имеют пигментов и поэтому не окрашены.
Прозрачность воды характеризуется максимальной глубиной, на которой еще виден диск Секки (диск, окрашенный в белый цвет, диаметром 20 см). Самые прозрачные воды в Саргассовом море – диск виден до глубины 66,5 м, в Тихом океане – 59 м, в мелких морях – 5…15 м, в реках 1…1,5 м, оз. Байкал – 40 м.
Нижняя (по глубине) граница фотосинтеза меняется также в зависимости от времени года, широты местности и др.
В темных глубинах океана в качестве источника зрительной информации организмы используют свет, испускаемый живыми существами за счёт процесса биолюминесценции. Химия процесса заключается в реакции окисления определённых органических соединений (люциферинов) при помощи белковых катализаторов. При этом избыточная энергия возбуждённых органических молекул выделяется в виде квантов света. Свет излучается импульсами в ответ на раздражения, поступающие из внешней среды. Биолюминесценция играет в основном сигнальную роль.
Специфические приспособления гидробионтов:
Из-за поглощения света в воде у гидробионтов слабо развита зрительная и лучше звуковая ориентация, т.к. звук распространяется в воде быстрее, чем в воздухе.
Для ориентации в глубине гидробионты используют гидростатическое давление толщи воды.
Китообразные используют эхолокацию на основе ультразвука.
Около 300 видов рыб способны генерировать электричество и использовать его для ориентации, сигнализации и защиты от врагов.
Наиболее древний способ ориентации водных животных – восприятие химического состава среды. Хеморецепторы их обладают чрезвычайно большой чувствительностью, что позволяет животным в тысячекилометровых миграциях по океану ориентироваться по запахам. Так, например, угри, кормящиеся в европейских реках и нерестящиеся у берегов Центральной Америки, реагируют на наличие этилового спирта концентрацией 1г на 6000 км 3 воды.
Некоторые гидробионты приспособились к фильтрационному способу питания, процеживая через себя большое количество воды с извлечением из неё необходимых веществ (пластинчато-жаберные моллюски и др.). Животные-фильтраторы играют огромную роль в биологической очистке водоёмов. Так, мидии, обитающие на площади дна в 1 м 2 , могут фильтровать до 150…280 м 3 воды за сутки, очищая её от взвешенных частиц.
Многие гидробионты адаптировались к условиям обезвоживания водоёмов (зарывание в ил, переход в состояние пониженной жизнедеятельности (гипобиоз) и др.).
Источник